Wolf-Ekkehard Lönnig
Mutationen: Das Gesetz der rekurrenten Variation
All competent biologists acknowledge the limited nature of the variation breeders can produce, although they do not like to discuss it much when grinding the evolutionary ax.
William R. Fix
Needless to say, I did not succeed in producing a higher category in a single step; but it must be kept in mind that neither have the Neo-Darwinians ever built up as much as the semblance of a new species by recombination of micromutations. In such well-studied organisms as Drosophila, in which numerous visible and, incidentally, small invisible mutations have been recombined, never has even the first step in the direction of a new species been accomplished, not to mention higher categories.
Richard B. Goldschmidt
Neue Arten sind experimentell weder durch die schrittweise Anhäufung von Genmutationen noch durch die Induzierung einzelner progressiver Mutationen hergestellt worden.
Werner Gottschalk
Mutations are merely hereditary fluctuations around a medium position…No matter how numerous they may be, mutations do not produce any kind of evolution.
Pierre-Paul Grassé
(On evolutionary novelties by chance mutations:) I have seen no evidence whatsoever that these changes can occur through the accumulation of gradual mutations.
Lynn Margulis
Mutations are a reality and while most of them are of no consequence or detrimental, one cannot deny that on occasion a beneficial mutation might occur [in relation to a certain environment, but usually not for a gene's function per se; Anmerkung von W.-E.L.; vgl. Diskussion]. However, to invoke strings of beneficial mutations that suffice to reshape one animal into the shape of another is not merely unreasonable, it is not science.
Christian Schwabe
I N H A L T S V E R Z E I C H N I S
Vorbemerkung: Als konstruktive Kritik
schlägt
ein Kollege vor, das Studium des
Beitrags zum leichteren Einstieg in
die
Thematik
mit dem Punkt
"c) Einleitung:
Die Ursprungsfrage" zu beginnen.
b) Hauptpunkte und Zusammenfassung
c) Einleitung: Die Ursprungsfrage
d) Die Antwort der heutigen Biologie: Mutationen
e) Mutationen - der alleinige Grund jeglicher Schöpfung?
f) Mutationen in der Pflanzenzüchtung
g) Die Ableitung des Gesetzes der rekurrenten Variation
h) Prognosen und Retrognosen auf Grund des Gesetzes der rekurrenten Variation
i) Die Ursachen des Gesetzes der rekurrenten Variation
j) Bestätigung durch Vavilovs Gesetz der Parallelvariation
l) Die generelle Bedeutung des Gesetzes der rekurrenten Variation
m) Der Darwinismus als Hindernis in der biologischen Forschung zur Zeit Mendels und heute
n) Einwände des Neodarwinismus zum Gesetz der rekurrenten Variation
o) Schlussbemerkungen zum Neodarwinismus
p) Alternativen
Wildarten, genetisches Potential und Parallelinduktion
Dr. G.s Kommentar zur Teleologie-Frage
Dr. H.: Was ist über die Geschichte des Gesetzes der rekurrenten Variation zu berichten?
EINWAND: Aber bedenken Sie doch die Jahrmillionen...
LINK: Evolution durch Genduplikationen?
q) Literatur
Weitere LINKS:
Utricularia (der Wasserschlauch)
Diskussion zu Utricularia Ursprung der Vogelfeder (vgl. auch die daran anschließenden Kapitel.)
a) Definitionen zum Begriff Naturgesetz
Keynote (Grundgedanke): "Wissenschaft ist
der Versuch, aus Einzelbeobachtungen etwas
allgemein Gültiges zu erschließen, aus Bekanntem auf Unbekanntes zu extrapolieren
und sich mit der Kritik gegenteiliger Argumente
auseinanderzusetzen."
Peter von Sengbusch
Zu diesem Aufsatz möchte ich einige Zitate zur Definition des Schlüsselbegriffs "Naturgesetz" vorausschicken, deren Bedeutung ich den Leser bitte, beim Studium der folgenden Ausführungen im Sinn zu behalten (Hervorhebungen im Schriftbild von mir):
"Naturgesetz, eine erfahrungsgemäß sich immer wieder bestätigende, aus dem Naturgeschehen abgeleitete Regel; im engeren Sinn ein allgemein anerkannter Lehrsatz einer Naturwissenschaft, d.h. eine Aussage, die bei Vorliegen gewisser experimentell prüfbarer Bedingungen Voraussagen über das Eintreten anderer Erscheinungen gestattet" (Brockhaus 1998, Bd. 15).
"Beim Vorliegen bestimmter experimentell prüfbarer Ausgangsbedingungen kann man auf Grund der Naturgesetze vorhersagen, welche neuen nachprüfbaren Verhältnisse sich einstellen werden (‡ Kausalität)" (Fischerlexikon 1976, Bd. 13).
"(Naturgesetze) sind nur durch Beobachten (Experiment) möglichst vieler ähnlicher Einzelfälle zu entdecken…sie zeigen einen Kausalzusammenhang und erlauben daher Vorausberechnung künftiger Ereignisse aus gegebenen Vorbedingungen" (Herder/Spektrum: Biologie; 1994, Bd. 6).
"Ein Prozeßgesetz enthält den Zeitfaktor. Es erlaubt die Prognose (oder Retrognose) zukünftiger (oder vergangener) Zustände eines Systems, falls die Werte der relevanten Variablen für wenigstens einen Zeitpunkt bekannt sind" (Hans Mohr, Herder/Spektrum: Biologie; 1994, Bd. 3, p. 178).
"Ein "Gesetz" ist eine gesicherte Aussage, die für eine Vielzahl von Systemen gilt." (Vom Verfasser kursiv) … Die Art, wie die Aussage gemacht wird, ob zum Beispiel mathematisch oder in einer natürlichen Sprache, ist dabei zweitrangig, falls den logischen und semantischen Ansprüchen der Wissenschaft Genüge getan ist" (Hans Mohr, Herder/Spektrum: Biologie; 1994, Bd. 3, p. 178).
"Im Falsifikationismus K. R. Poppers sind Naturgesetze gegen Widerlegungsversuche erfolgreiche Sätze" (Brockhaus 1998).
Zur Historie des Begriffs Naturgesetz vgl. die erste Fußnote (#) und zu den wesentlich weniger problematischen Begriffen Mutation und Variation konsultiere der mit diesen Termini noch nicht vertraute Leser bitte die einschlägigen Nachschlagewerke und Lehrbücher (oder siehe z.B. Punktmutationen und Wirkung der Punktmutationen, sowie zu den Genduplikationen und Polyploidien den im Inhaltsverzeichnis schon aufgeführten Link).
b) Hauptpunkte und Zusammenfassung: Nach Aussage der neodarwinistischen Evolutionstheorie sind
Mutationen der "einzige und alleinige Grund jeglicher Neuerung und Schöpfung in der belebten Natur"
(Jacques Monod). Auf der DNA-Ebene ist dazu der Ursprung von mehr als 5000 verschiedenen (nicht voneinander
ableitbaren) Genfamilien mit über 50000 Genen zu erklären. Dabei wird der Selektionstheoretiker
hinzufügen, dass jede mutative Veränderung mit Selektionsvorteilen einhergehen muss, wenn sie sich
in einer Population durchsetzen soll. Diese seit Jahrzehnten als Lehrsatz geltende Behauptung wird hier in Frage
gestellt: Kann der Mechanismus der Mutation von Erbinformation - im Zusammenhang mit Rekombination und Selektion -
tatsächlich jegliche Schöpfung in der belebten Natur erklären? Im folgenden Aufsatz wird die These
begründet, dass keines der uns bisher bekannt gewordenen Mutationsereignisse den nach Monod zitierten
Allerklärungsanspruch zum Ursprung der Lebensformen rechtfertigt. Die mutativ erzeugten
Phänotypabweichungen folgen vielmehr dem Gesetz der rekurrenten Variation ("bei allen
hinreichend umfangreichen Mutageneseexperimenten mit Eukaryonten entstehen Mutanten in einem (meist sehr)
großen, jedoch begrenztem oder asymptotisch gegen Null laufenden Spektrum der Phänotypen" -
vgl. erste Prognose unten) und weisen damit auf mehr oder weniger weite Art-, Evolutions- und
Züchtungsgrenzen hin.
Abgesichert wird das Gesetz der rekurrenten Variation vor allem durch die Saturation mutagenesis ('Sättigungsmutagenese'): Diese zum Nobelpreis (1995) führende, erfolgreiche Methode des Studiums der Gene, war und ist nur möglich, weil das Genom in bezug auf bestimmte Fragestellungen (ob, wie ursprünglich, "screen for embryonic lethals" mit annähernd 18 000 Mutationen, oder bei Funktionsuntersuchungen bestimmter Bereiche von Genen, Proteinen oder neuerdings sogar ganzer Genome) auch zu "sättigen" ist, anstatt unendlich viele neue funktionsfähige Sequenzen mit neuen Phänotypen zu generieren. Die Möglichkeiten zur Bildung funktionsfähiger Allele mit dem Resultat abweichender Phänotypen sind eindeutig begrenzt und die Mehrzahl dieser sich phänotypisch auswirkenden mutanten Allele zeichnet sich durch ihren Defektcharakter aus (dazu 3 Beispiele: "…in the inherited recessive deseases cystic fibrosis and phenylketonuria, more than 200 different defective alleles of each gene have been identified in studies of affected children throughout the world"1a - und "…at least 95% of all amino acid substitutions, i.e., nonsynonymous mutations, in Hbb are functionally unacceptable in homozygous state1b - von den zahlreichen größeren Sequenzabweichungen ganz zu schweigen). Interessanterweise wird in dem Nobel Press Release von 9. Oktober 1995 "to damage" mit "to mutate" gleichgesetzt. In Übereinstimmung mit dieser Gleichsetzung gibt es keinerlei Befunde – nicht einmal einen erfolgversprechenden Ansatz - für die Bildung neuer in der Natur beständiger Arten durch experimentelle Mutagenese, geschweige denn einer saturation mutagenesis etwa für eine bereits erfolgte Bildung ganzer Serien neuer Arten. Der Kontrast zwischen dem neodarwinistischen Postulat mutativer Artbildung und den experimental-biologischen Realitäten wird vielleicht noch deutlicher, wenn man sich vor Augen hält, dass die mutative Artbildung letztlich unbegrenzt sein soll (Entstehung sämtlicher Spezies der Erdgeschichte bis in die Gegenwart durch Mutation und Selektion).
Zur Falsifikationsfrage ist (daher) festzustellen, dass im Prinzip das Gesetz der rekurrenten Variation durch die mutative Neubildung primärer Arten (durch den Aufbau neuer Gene, Genwirkketten und Genfamilien zur Bildung neuer anatomischer, physiologischer und ethologischer Strukturen und Systeme sowie des Aufbaus genetisch-plasmatischer Barrieren [vgl. ARTBEGRIFF] als Gesetz widerlegt werden könnte. Es würde jedoch selbst in diesem äußerst unwahrscheinlichen Falle in der Form einer jederzeit testbaren und reproduzierbaren Regel von weitestreichender Gültigkeit bestehen bleiben - vergleichbar etwa mit dem Status der Mendelschen Gesetze, von denen es bekanntlich einige Ausnahmen gibt.
Für solche bisher völlig unbekannten Fälle wäre zunächst nur der äußerst geringe Teil der Strukturaufbau-Mutationen zur Bildung neuer funktionaler DNA-Sequenzen von primärem artbildungstheoretischem Interesse, d.h. der Teil, der über die Regel der rekurrenten Variation hinausgehen würde (die zweite Frage wäre die nach dem Aufbau der plasmatischen Barriere). Zur Klarstellung der Kernfragen müsste dann jeweils deutlich zwischen dem Spektrum rekurrenter Phänotypabweichungen und dem Phänotyp aufgrund der Neubildung von Sequenzen sowie der Entstehung der genetisch-plasmatischen Barriere unterschieden werden. Die Artbildungsfrage liegt - im Gegensatz zu den von der Synthetischen Evolutionstheorie bisher zitierten Beispielen - jenseits der rekurrenten Variation.
Es sei jedoch in diesem Zusammenhang besonders betont, dass bislang nicht einmal - in der Natur beständige - sekundäre Arten (d.h. durch Strukturabbau entstandene Arten) mutativ erzeugt werden konnten:
In der Pflanzengenetik sind in Zehntausenden von mutationsgenetischen Experimenten Millionen und Abermillionen von Mutationen induziert worden sind. Bis heute ist mir kein einziges Beispiel bekannt, dass dadurch in der Natur beständige neue Pflanzenarten entstanden wären. Und das gleiche trifft auf die Frage nach der Artbildung bei Drosophila und anderen Organismen zu. Auch meine eigenen umfangreichen mutationsgenetischen Untersuchungen an Pflanzen sind mit dieser Aussage in voller Übereinstimmung. Wenn es hingegen solche Beispiele gäbe, dann dürfen wir davon überzeugt sein, dass sie praktisch in jedem Lehrbuch der Biologie aufgeführt wären.
Die gesamte Gentechnologie mit ihrem Unternehmen, spezifische DNA-Sequenzen mit spezifischen Funktionen von einem Organismus auf einen (meist) andersartigen zu übertragen, ist zugleich ein beredtes Zeugnis davon, dass die uns bekannten (definitionsgemäß richtungslosen) Mutationserscheinungen nicht ausreichen, um solche spezifischen Sequenzen zu erzeugen.
Darüber hinaus zeigt der folgende Aufsatz, inwiefern der weltweite Zusammenbruch der "Mutationszüchtung" als eigenständige Pflanzenzuchtmethode in den achtziger Jahren des letzten Jahrhunderts die Ableitung des Gesetzes der rekurrenten Variation ermöglicht und bestätigt hat.
Das Gesetz trifft auf alle Eukaryonten zu. Bakterien hingegen können aufgrund des regulären artübergreifenden horizontalen Gentransfers durch Plasmide und Bakteriophagen eine Sonderstellung in dieser Frage einnehmen (Transformation, Transduktion).
c) Einleitung: Die Ursprungsfrage
Angesichts der komplexen Ordnung und der unermesslichen Vielfalt der Organismen erhebt sich unausweichlich die Frage nach dem Ursprung dieser Ordnung: Wie ist diese genial-komplexe Vielfalt der Lebensformen und -funktionen entstanden? Welche Gesetzmäßigkeiten können wir erkennen? Lässt sich der Ursprung aller Lebensformen auf physikochemische Prozesse zurückführen? Wie steht es mit den in naturwissenschaftlichen Kreisen weithin verbotenen, ja häufig sogar mit großer Polemik zurückgewiesenen Fragen: Spielen hier zielgerichtete Faktoren eine Rolle, oder vielleicht sogar die Rolle? Ist der genetische Code samt tausender spezifischer DNA-Sequenzen programmiert entstanden? Stehen hinter dem Ursprung der genialen Ordnung des Lebens Intelligenz, Weisheit, Geist und Gott? "Teleologisch-finalistische Interpretationen sind bekanntlich naturwissenschaftlich nicht erlaubt" bemerkt L.-C. Schulz.1c Dieses Denkverbot sollte uns anspornen, der Sache um so genauer auf den Grund zu gehen.
d) Die Antwort der heutigen Biologie: Mutationen
"Nach Jahrmilliarden blinder Mutationen, die gegen die nachgebenden Grenzen der Umwelt anstürmten, entstand aus den Mikroben der Mensch" behauptet H. J. Muller2, der für seine Arbeiten auf dem Gebiet der Mutationsgenetik den Nobelpreis erhielt. Der Mensch war bei diesem Mutationsgeschehen weder geplant noch überhaupt in irgendeiner Weise vorherzusehen. Wenn man das ganze postulierte Evolutionsdrama noch einmal von vorn durchspielen könnte, dann "würden die Fauna und die Flora der Erde radikal anders aussehen, und die Art Mensch - dieses "unwahrscheinliche und zerbrechliche Wesen" - würde vermutlich nicht existieren"3. In diesem Punkt sind sich die meisten Biologen einig. Der Homo sapiens "weiß nun, dass er seinen Platz wie ein Zigeuner am Rande des Universums hat, das für seine Musik taub ist und gleichgültig gegen seine Hoffnungen, Leiden oder Verbrechen"4, dessen Glückszahl zufällig einmal auftrat. Es gibt keinen rational fassbaren Plan, kein Ziel, keinen Sinn mehr im vierdimensional/gekrümmten Universum.
In der 1991er und 1998er Auflage von Strasburgers Lehrbuch der Botanik schreibt P. Sitte: "Der Selektion als restriktivem Prinzip stehen zufällige Erbänderungen (Mutation, Rekombination) gegenüber. Ihnen sind der unermessliche Artenreichtum und die vielen physiologischen, ökologischen und eben auch morphologischen Problemlösungen in der Organismenwelt zuzuschreiben.''5 Mutationen "sind die Grundlage jeder Evolution", schreibt F. Ehrendorfer in denselben Ausgaben des Lehrbuchs6 im Einklang mit anderen führenden Biologen:7 Ohne Mutationen wäre unsere Erde so tot wie die Oberfläche des Mondes, käme die Evolution letztlich zum Stillstand beziehungsweise hätte erst gar nicht eintreten können, gäbe es keine naturgesetzliche Erklärung des Lebens, keine Vergangenheit und keine Zukunft des Lebens - Mutationen sind der "einzige und alleinige Grund jeglicher Neuerung und Schöpfung in der belebten Natur"8, sie sind eine zentrale Grundlage der modernen Biologie. - Wobei der Selektionstheoretiker hinzufügen wird, dass jede Veränderung mit Selektionsvorteilen einhergehen muss, wenn sie sich durchsetzen soll.
e) Mutationen - der alleinige Grund jeglicher Schöpfung?
Aufgrund eines reichen Erfahrungsschatzes aus Mutationsexperimenten mit Hunderttausenden von Pflanzen und zwei umfangreichen Mutantensortimenten (sowie der systematischen Analyse der mutationsgenetischen Literatur zur Pflanzen- und Tierwelt) lautet meine Antwort eindeutig, dass keines der uns bisher bekannt gewordenen Mutationsereignisse die oben zitierte Allerklärungsansprüche zum Ursprung der Lebensformen rechtfertigt.9 Die Ansprüche liegen vielmehr "jenseits" aller Erfahrung und theoretischen Forschungsergebnisse und sind weitgehend eine Deduktion aus dem anfangs zitierten Grundsatz, dass, "finalistisch-teleologische Interpretationen naturwissenschaftlich bekanntlich nicht erlaubt" sind: Da ein zielgerichteter Aufbau von DNA-Sequenzen (und überhaupt jeglicher Strukturen des Lebens) grundsätzlich entfällt und die vielfach beobachteten Zufallsmutationen die einzige bisher bekannte naturwissenschaftliche Antwort auf die Frage nach dauerhaften Veränderungen bei den verschiedensten Lebensformen sind (von einigen Randerscheinungen wie dem horizontalen Gentransfer bei Bakterien und der zur Zeit wieder stärker betonten Endosymbiontenfrage10 einmal abgesehen) müssen sie auch die ihnen oben zugeschriebene Rolle gespielt haben. Wie noch weiter zu begründen ist, liegt hier eine ungerechtfertigte Extrapolation vor, die in wesentlichen Punkten der Schlussfolgerung eines Beobachters gleicht, der von den einzigen ihm bisher bekannten Veränderungen an Automobilen durch Verkehrsunfälle auf den Modus ihrer Entstehung schließt.
Im Rahmen dieses Beitrags ist nun nicht beabsichtigt, die Fülle der fachspezifischen (klassischen und molekularbiologischen) Details der verschiedenen bisher festgestellten Mutationsereignisse aufzuführen.11 Vielmehr wollen wir hier anhand einiger Beispiele vom Phänomen ausgehen und die Frage stellen, ob zunächst einmal allein von dieser Ebene aus gewisse Gesetzmäßigkeiten zu beobachten sind, die eine klar nachvollziehbare Antwort auf die Frage nach der Leistungsfähigkeit der Mutationen geben.
f) Mutationen in der Pflanzenzüchtung
Als die Synthetische Evolutionstheorie Ende der dreißiger Jahre beanspruchte, mit ihrem Zwei-Faktorensystem von Mutation und Selektion die wissenschaftlich verbindliche Antwort auf die Frage nach der Entstehung aller Lebensformen zu geben, rechneten viele Genetiker und Pflanzenzüchter mit einer weltweiten Revolution in der Pflanzenzüchtung: Wenn die Mutationen das Rohmaterial für die Entstehung aller Gene und Proteine, aller physiologischen Prozesse und anatomischen Strukturen sämtlicher Lebensformen geliefert hatten, dann mussten in Anwendung dieser Faktoren auch die erstaunlichsten Erfolge in der Züchtungsforschung für möglich gehalten werden. Drei Zeitraffer hatte man in der Hand: Vervielfachung der Mutationsraten, gezielte Rekombination und intelligente Selektion. Mit einem an Euphorie grenzenden Optimismus wurde in den USA sowie in mehreren Ländern Europas und Asiens das neue Fach Mutationszüchtung in Angriff genommen. Simmonds schreibt dazu in seinen Principles of Crop Improvement:
Earlier overoptimism, to the effect that induced mutations were about to revolutionize plant breeding, has given place to a more sober appreciation of the technique as a valuable supplement to more conventional techniques in certain, rather restricted circumstances.....very many programmes failed, especially in the early days of overoptimism, to produce anything useful because they were not fulfilled. Nowadays we see mutation-induction simply as one technique which is occasionally useful in enlarging the genetic base of a programme in a limited and highly specific fashion.11a
Weiter beschreibt F. Leibenguth in seiner Züchtungsgenetik das Gesamtresultat nach etwa vierzig Jahren intensivster Forschung wie folgt:
Die allermeisten Mutanten zeichnen sich durch einen negativen Selektionswert aus. Nach den Erfahrungen an Getreidearten und Leguminosen beträgt der Anteil züchterisch brauchbarer Mutanten 0,5 bis 1 Prozent der in den Versuchen selektierten Genotypen. Hinzu kommt häufig ein negativer Effekt auf andere Komponenten des pleiotropen Merkmalsspektrums, der den züchterischen Wert einer positiven Mutante wieder herabsetzt. Daraus geht hervor, daß das ursprüngliche Ansinnen, die zeitraubenden und kostspieligen Methoden der Rekombinationszüchtung durch, "Mutationszüchtung" zu ersetzen, heute nicht mehr aktuell ist. Man betrachtet die Mutationszüchtung weniger als selbständige Zuchtmethode, sondern zieht sie vielmehr zur Ergänzung der traditionellen Methoden heran (Hervorhebung von mir).12
Leibenguth bemerkt außerdem, dass die Mutationszüchtung für die Tierzucht überhaupt nicht zu gebrauchen ist. "Denn Tiere sind im Unterschied zu Pflanzen genetisch schärfer ausbalanciert; daher wirken bei ihnen alle Arten von Mutationen noch häufiger letal und stärker vitalitäts- und fertilitätsmindernd." Hier hat die Mutationszüchtung keinerlei Zukunft.
In der Pflanzenzüchtung sind weniger als ein Prozent aller induzierten Mutanten für Feldversuche als geeignet erachtet worden.13 Davon haben wiederum nur etwa 0,5 bis 1 Prozent die weiteren Untersuchungen bis zum kommerziellen Gebrauch bestanden. In der Pflanzenzüchtung ist daher das Verhältnis von negativen beziehungsweise unbrauchbaren zu positiven Mutanten geringer als 10.000:1. Rechnet man mit 0,5 Prozent aller induzierten Mutanten für weitere Untersuchungen und davon wiederum mit 0,5 Prozent mit positivem Selektionswert, so ist das Verhältnis schon 40.000:1. Ein geschätzter Mittelwert um 25.000:1 dürfte damit nicht unrealistisch sein. Bei den genetisch schärfer ausbalancierten Tieren ist die Situation schon so schwierig, dass nicht einmal mehr genaue Zahlen zur Ermittlung eines realistischen Verhältniswertes zugrunde gelegt werden können. Wenn wir für ein Selektionsziel nur eine Zehnerpotenz höher gehen, kommen wir schon zu einem Verhältnis von 100.000 bis 400.000 negativen (bzw. 'neutralen') Mutanten zu einer positiven.
Aufgrund solcher Daten und Erfahrungen haben die meisten kommerziellen Zuchtbetriebe die Mutationszüchtung ganz aus ihrem Programm gestrichen.14
"One can only conclude that the results from mutation breeding in varietal development of the major field crops have been rather meager in relation to the efforts expended." So fasst J. M. Poehlmann das Gesamtresultat der Mutationszüchtung zusammen.14a
"Der Versuch durch Bestrahlung ertragreichere Sorten zu züchten, erwies sich trotz eines großen finanziellen Aufwands weitgehend als Fehlschlag" (Peter von Sengbusch).14b
"Das Leitziel der praktischen Pflanzenzüchtung, mit Hilfe der Mutationsauslösung neue Möglichkeiten einer schrittweisen und stetig fortgesetzten Verbesserung bewährter Zuchtsorten zu erschließen, konnte...nicht verwirklicht werden" (Fischbeck, Röbbelen und Stutzer). Und speziell zu dem neodarwinistischen Konzept der "Mikromutationen" bemerken die Autoren weiter: "Auch das abgewandelte Konzept einer direkten züchterischen Nutzung sogenannter "Mikromutationen" blieb ohne Erfolg, weil die damit erzielbaren Zuchtfortschritte deutlich hinter der züchterisch nutzbaren Variabilität zurückblieben, die sich aus dem breiten Strom konventioneller Kombinationszüchtung entwickeln ließ".14c
Die nach den Prämissen der Synthetischen Evolutionstheorie erhoffte und fest erwartete weltweite Revolution in der Pflanzenzüchtung hat sich als Fehlschlag erwiesen. Die Mutationen leisten nicht das, was die Neodarwinisten von ihnen behaupten. Die wenigen brauchbaren Mutationsereignisse in der Pflanzenzüchtung beruhen hauptsächlich auf dem Abbau von Strukturen und Funktionen (alkaloidfreie Lupinen, Raps ohne Erucasäure, Erbsen ohne Fiederblätter etc.). Mit dem Abbau von Strukturen kann man jedoch nicht den Aufbau der gesamten Organismenwelt erklären.
Wenn auch für die Pflanzenzüchtung auf der Grundlage des Neodarwinismus der erhoffte Erfolg ausgeblieben ist, so wurde doch durch die Mutationszüchtung das wissenschaftliche Interesse "mit einer raschen Ausweitung der Kenntnisse über die Lokalisierung genetischer Effekte im Genom wichtiger Kulturpflanzen belohnt."15 Das heißt, die genetische Grundlagenforschung hat von diesem Unternehmen reichlich profitiert - oder in den Worten von Sengbuschs: "Obwohl sich die Produktion pflanzlicher Mutanten wirtschaftlich nicht rentiert hat, erwies sie sich als das wohl wirkungsvollste experimentelle Instrument moderner Grundlagenforschung."15a Das für unsere Fragen nach dem Ursprung der Lebensformen wichtigste Ergebnis für die Grundlagenforschung bezeichne ich als Gesetz der rekurrenten Variation,mit dem wir uns jetzt näher beschäftigen wollen.
g) Die Ableitung des Gesetzes der rekurrenten Variation
Ein wesentlicher Punkt, der die Schlussfolgerung von der Unwahrscheinlichkeit des Ursprungs der Lebensformen durch Mutation und Rekombination unterstützt, ist die seit Jahrzehnten bekannte und an den verschiedensten Organismen immer wieder gemachte Erfahrung, dass sich das Mutantenspektrum nach mehrfach wiederholter mutagener Behandlung verschiedener Linien oder Arten nur noch geringfügig vergrößert. Das heißt, es treten immer wieder die gleichen Mutanten auf. Der Genetiker Hans Stubbe hat diesen Punkt für seine Studien am Löwenmäulchen (Antirrhinum majus L.) nach 39 Jahren intensiver Mutationsforschung wie folgt zusammengefasst:
Die immer bessere Kenntnis der Mutanten von Antirrhinum hat einige wesentliche Erfahrungen gebracht. Mit jedem neuen großen Mutationsversuch ergab sich im Laufe der Jahre, daß die Zahl der wirklich neuen, erstmalig erkannten Mutanten immer geringer wurde, daß also die Mehrzahl der auftretenden erblichen Änderungen schon bekannt war.16
Ähnlich stellt ein erfahrener Genetiker wie Werner Gottschalk fest:
Je größer die Sortimente sind, um so schwieriger ist es, sie durch neue Mutationstypen zu erweitern. Es entstehen hierbei bevorzugt Mutanten, die bereits existieren.17
Anders formuliert, strebt die Zahl der neuen Mutantentypen mit immer weiteren großen Mutationsversuchen asymptotisch gegen Null.18 Die Ergebnisse von U. Lundqvist aus ihrer langjährigen Erfahrung mit der Mutationszüchtung der Gerste veranschaulichen die rekurrent auftretenden Mutanten in hervorragender Weise. Sie stellte fest, dass in den letzten fünfzig Jahren in Svalöf (Schweden) etwa 9000 Gerstenmutanten isoliert worden sind.19 Die Letalmutationen mitgerechnet wurden allein in diesem Zeitraum mindestens 100 000 Mutanten festgestellt.20
Aus der Arbeit von Lundqvist seien einige Beispiele für das wiederholte Auftreten bestimmter Mutantentypen zitiert (Tab.1).
Tabelle 1: Beispiele für das wiederholte Auftreten bestimmter Mutantentypen der Gerste nach Lundqvist (deutsche Übersetzung von mir)
|
Mutante |
wiederholt aufgetreten |
Zahl der Genloci |
|
Erectoides (dichte Ähren) |
205 mal |
26 |
|
Praematurum (frühe Reife) |
110 mal |
9 |
|
Eceriferum (wachslos) |
1527 mal |
76 |
|
Breviaristatum (kurze Grannen) |
140 mal |
17 |
|
Exrubrum (ohne Anthozyan) |
61 mal |
18 |
|
Macrolepis (deckspelzenähnliche Hüllspelzen) |
40 mal |
1 |
|
Hexastichon (sechszeilig) |
41 mal |
1 |
|
Intermedium (zwischen zwei- und sechszeilig) |
144 mal |
11 |
|
Powdery mildew resistent (mehltauresistent) |
154 mal |
? |
Das heißt, dass allein diese neun Mutanten alles in allem 2422 mal aufgetreten sind. Nach rund 50 jährigen Mutagenese-Experimenten verteilten sich im Jahre 1988 die 9000 Gerstenmutanten auf genau 93 Mutantentypen oder -klassen (U. Lundqvist). (Details vgl. Artbegriff, S. 547; ich habe jedoch noch nicht herausfinden können, ob in den letzten Jahren weitere Typen bzw. Gruppen dazu gekommen sind). Viele dieser Mutanten sind über 100mal aufgetreten und manche sogar über 1000mal. Wie bei der Gerste, so liegen auch zum Beispiel für die Erbse die gleichen Erfahrungen an verschiedenen Institutionen mehrerer Kontinente vor (weitere Tabellen vgl. Lönnig 1993). Andere Hauptträger der Welternährung wie Reis, Mais und Soja dürften der Gerste kaum mehr nachstehen (selbst der allotetraploide Hartweizen und der allohexaploide Brotweizen sind mutationsgenetisch intensiv bearbeitet worden). - Von den in allen Forschungsinstituten zusammengenommen wahrscheinlich in die Millionen gehenden Gerstenmutanten sind bislang 25 in die Liste der Gerstenkultivare eingegangen und 33 haben über Rekombination Eingang in den kommerziellen Anbau gefunden.21 Die meisten dieser Linien werden jedoch inzwischen nicht mehr angebaut. Obwohl "Mutanten für praktisch jedes erkennbare Merkmal existieren"22, ist für die Züchtungsforschung bislang nicht mehr dabei herausgekommen. Es sei in diesem Zusammenhang besonders betont, dass etwa von der Bildung neuer in der Natur beständiger Arten nirgends die Rede ist. Zu den rekurrent auftretenden Mutanten schrieben H. Kuckuck und A. Mudra schon 1950:
Wie umfangreiche Versuche besonders bei Gerste gezeigt haben, können durch Röntgenbestrahlung wohl sämtliche Formen des Weltsortiments mutativ erzeugt werden (…). Diese erzeugten Mutanten haben sich teilweise mit den gleichen Formen des Weltsortiments als genetisch identisch erwiesen. In anderen Fällen werden gleiche Phänotypen durch verschiedene Gene bedingt; es liegen dann sogenannte heterogene Merkmalsgruppen vor.23
Ähnliche Beobachtungen gibt es auch für andere Kulturpflanzen wie Reis und Mais. Neue Arten sind dabei niemals entstanden! Statt der Bildung neuer Formen und Arten wiederholt sich ununterbrochen das gleiche Mutantenspektrum, so dass die Methode auch für die Züchtungsforschung in der Regel schon nach wenigen Versuchen nichts mehr bringt. Hatte man Ende der sechziger Jahre beispielsweise noch geglaubt, mit Hilfe der Mutationszüchtung Getreideproteine verbessern zu können, so stellen Micke und Weindl nach rund eineinhalb Jahrzehnten intensivster Anstrengungen auf diesem Gebiet zusammenfassend fest:
Our programme on the improvement of grain protein has now come to an end ... during the 14 years of the programme it had to be recognized that the matter is more complicated and that there are some mutual limitations of quantity and quality!24
Selbst die massivste Mutationsinduktion ändert nichts an der Tatsache des Selektionslimits. Dasselbe ist für das Tierreich wiederholt festgestellt worden (ausführliche Diskussion der Selektionslimits in der Artbegriffsarbeit).25 Zusammenfassend sei mit D. L. Hartl einer der hervorragendsten zeitgenössischen Populationsgenetiker zu dieser Frage zitiert:
Progress under artificial selection cannot go on forever, of course. As noted earlier, the population will eventually reach a selection limit, or plateau, after which it will no longer respond to selection. ...However, many experimental populations that have reached a selection limit readily respond to reverse selection. (Hervorhebung im Schriftbild von mir).26
Und einige Jahre darauf betonen Hartl und Jones wiederum:
Population improvement by means of artificial selection cannot continue indefinitely. A population may respond to selection until its mean is many standard deviations different from the mean of the original population, but eventually the population reaches a selection limit at which successive generations show no further improvement.26a
Sowohl im Pflanzen- als auch im Tierreich ist immer wieder festzustellen, dass auch bei fortgesetzt intensiver Mutationsinduktion Selektionsgrenzen erreicht werden, die praktisch nicht mehr überschritten werden können. Das Spektrum der Mutanten ist zwar groß, aber begrenzt! Es ist überdies bei entsprechenden Populationsgrößen durch die rekurrent auftretenden Mutanten beliebig reproduzierbar, so dass wir das Gesetz der rekurrenten Variation wie folgt formulieren können:
Beim spontanen Mutationsprozess in der Natur entsteht bei gleichen genetischen Voraussetzungen grundsätzlich das gleiche Mutantenspektrum, aber wegen der geringeren Mutationsfrequenz sind hier wesentlich größere Populationen und Zeiträume erforderlich.
Das Gesetz gilt auch für heterozygote Beispiele ("Fremdbefruchter"), nur dass durch Genzufuhr bei Kreuzungen die Kurven entsprechend langsamer abfallen. - Bakterien können hingegen einen Sonderstatus durch ihren artübergreifenden horizontalen Gentransfer (permanente Zufuhr neuen genetischen Materials) aufweisen (natürliche Transformation und Transduktion).
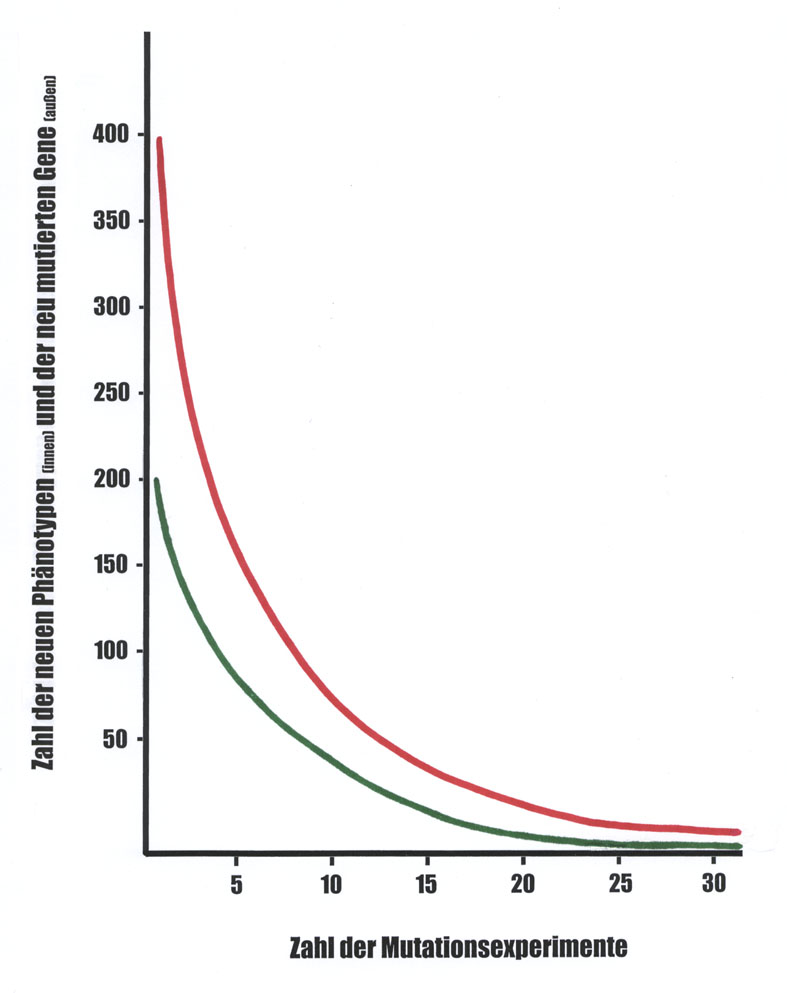
Abb. 1: Die Abbildung veranschaulicht die Abnahme neu auftretender Mutanten mit zunehmender Zahl der durchgeführten Experimente. Abszisse: Zahl der Mutationsexperimente. Ordinate: Zahl der neuen Phänotypen (grüne Kurve) beziehungsweise Zahl der neu mutierten Genloci (rote Kurve). Da Mutationen in verschiedenen Loci zu ähnlichen Phänotypveränderungen führen können, beginnt die Kurve der neuen Phänotypen tiefer und nähert sich der Abszisse schneller als die der rekurrent mutierten Loci. Das Redundanzproblem [siehe unten] läßt die Distanz zwischen beiden Kurven in vielen Fällen noch größer werden. Generell fallen für verschiedene Organismen die realen Kurven unterschiedlich aus. Der genaue Kurvenverlauf hängt weiter vom Umfang und von der Art der Experimente ab: So wurden in letzter Zeit zum Beispiel die meisten Gene von Arabidopsis mit Transposons markiert und damit in der Regel auch mutiert (ZIGIA-Projekt). Würde man mutierte Gene und veränderte Phänotypen dieses Projekts graphisch im Prinzip wie oben darstellen und mit entsprechend dargestellten Ergebnissen anderer Mutagenesemethoden vergleichen, so wären abweichende Kurven zu erwarten (Kurvenhöhen und -längen). Allen gemeinsam ist jedoch die Endlichkeit des Mutantenspektrums (vom mikro-quantitativen Rest [siehe unten] einmal abgesehen).
Überdies ist anzumerken, dass auch im Falle einer weiteren (wenn auch
zunehmend absinkenden) Phänotypbildung in asymptotischer Näherung an die Abszisse (siehe
Abb. 1) die Evolution nicht grenzenlos ist. Diesen Punkt hat D. Einhorn [war Professor in Wien] einmal sehr
überzeugend diskutiert. Zu seinen folgenden Passagen ist jedoch vorauszuschicken, dass
Einhorn die gesamte Variabilität der Arten im Sinne hatte, d.h. inklusive der
Mendelschen Rekombination und nicht (nur) die Bildung neuer Phänotypen durch Mutationen (soweit
damals bekannt). Der
Unterschied zwischen diesen beiden Ebenen der Betrachtung wird sofort klar, wenn man sich
vergegenwärtigt, dass allein bei 30 (dreißig)
heterozygoten Allelpaaren## im
dominant-rezessiven Erbgang schon das Potential für 1.073.741.824 (mehr als
eine Milliarde!)
verschiedene homozygote Rekombinanten vorliegt (vgl. weiter ARTBEGRIFF und Punkt 8) unten). Das von
Einhorn geschilderte Prinzip trifft dann natürlich auch auf die sehr viel geringere und
laufend weiter absinkende Bildung mikro-quantitativer Phänotypen im Laufe fortgesetzter
Mutagenese-Experimente zu. In diesem Sinne möchten wir uns seine folgenden Ausführungen näher
ansehen (Schriftbild von mir):
Man behauptet sehr häufig von deszendenztheoretischer, speziell
Darwin-Haeckelscher Seite,
daß die Grenzen der Variabilität im Bereich der Erfahrung ungemein w e i t gezogen
sind. Das
ist aber prinzipiell falsch, denn die Variabilität erscheint uns in der Erfahrung gar nicht grenzenlos,
u
n g e m e i n w e i t, sondern lediglich u n g e m e i n r e i c h,
doch bei allem
tatsächlich vorhandenen ungeheuren Reichtum der Variationen hat noch niemand etwa eine Säugetierform
oder eine Vogelspezies im Bereiche der Variationen eines Amphibiums gesehen und geglaubt - und viel weniger
können wir uns vorstellen, daß trotz noch so mühe- und kunstvoller Einwirkung von außen
nicht wieder ein anderer Moner, sondern ein Wirbeltier im Bereiche der Varianten eines Moneren zu finden
wäre. Die prinzipielle Verwechselung des ungeheuren Reichtums der Variabilität mit der
unermeßlichen Weite derselben ist die Achillesferse, das "Proton Pseudos" der Darwin-Haeckelschen
Deszendenztheorie. Nur wegen dieses fast unerschöpflichen Reichtums, nicht aber der Weite, ist es
nicht möglich, auf diesem Wege der morphologischen und physiologischen Artabgrenzung zu einer konstanten,
nicht mehr variablen organischen Größe vorzudringen, nur wegen der beinahe unbeschreiblichen Fülle
der Varianten wissen wir nicht, wo wir im Bereiche unserer Erfahrung die letzten festen Grenzen der
Variabilität für ein gegebenes Wesen abzustecken haben. Diese Schwierigkeit aber, die Grenzen zu
bestimmem, dieses Nichtwissen um die letzten Grenzen eines Organismus verwandelt sich sofort der Deszendenztheorie
in ein sicheres Wissen um die Grenzenlosigkeit der Variabilität.
Ein Beispiel aus dem Gebiete der Mathematik kann die Sache noch besser veranschaulichen und
erklären: Zwischen 1 und 2 liegt eine unfaßbare Fülle und Mannigfaltigkeit von Werten und Formen,
wie 1 1/2, 1 1/3, 1 1/4,...1 1/100,...1 2/100 000 usw. ins Unendliche, also eine Unendlichkeit im kleinen,
die aber immer in den unverrückbaren Grenzen von 1 und 2 eingeschlossen verbleibt und nie zu 3, 4, zur
Unendlichkeit im großen werden kann. Nun stellen wir uns vor, daß wir etwa vom Werte 1 1/2 ausgehen,
die einzelnen verschiedensten kleinen und verschwindend kleinsten Werte und Formen genau sichten und mustern,
sowohl in der einen Richtung als auch in der anderen uns bewegen und uns durchaus bemühen, zu den letzten
Grenzen, zu 1 und 2 vorzudringen, so werden wir gewiß eine unbeschreibliche Fülle von Werten und Formen
kennen lernen, ohne doch die letzten Grenzen selber feststellen zu können; ja, je mehr Unterschiede wir
beachten werden und je genauer, ein um so größerer Reichtum wird sich unseren Augen offenbaren, um so
weiter werden wir von den letzten Grenzen entfernt sein, um so mehr werden wir geneigt sein zu glauben,
daß es überhaupt keine Grenze gebe - denn das ist die eigentliche Natur der Unendlichkeit im kleinen,
daß, je mehr Zwischenstufen wir kennen, je präziser, um so mehr noch zu erkennen bleibt, um so weiter
von uns die letzte Grenze zu rücken scheint -, und wir dürfen doch wohl nicht ernstlich glauben,
daß wir darum bereits längst über alle Grenzen hinauskommen und uns im anderen Grenzenlosen, im
Unendlichen im großen befinden. So ist es auch mit der Variabilität. Daraus, daß wir
eine gewaltige Formenfülle eines bestimmten organischen Wesens erleben können, folgt noch keineswegs,
daß die Variabilität dieses Wesens bereits gar keine Grenzen habe, daß sie in
progressiver Richtung grenzenlos, absolut unendlich sei, daß wir im Umkreis der unendlichen Varianten etwa
eines Marsupialiers (der unendlichen Varianten zwischen 1 und 2 auch tatsächlich die unzähligen
Varianten einer Fledermaus (der unzähligen Varianten zwischen "5 und 6", 5 1/2, 5 1/3...), eines Wales
usw. finden könnten, daß es für diese Variabilität überhaupt gar keine Grenzen gebe,
daß wir von der Unendlichkeit im kleinen (1 und 2) aus wirklich die ganze Unendlichkeit aller organischen
Formen, die Unendlichkeit im großen (1 - unendlich) hervorbringen könnten. Diesen Schluß zieht
nun aber die Deszendenztheorie.26b(1) Die oben erwähnte asymptotische Näherung der Zahl neuer
phänotypisch unterscheidbarer Mutanten an die Abszisse (Abb. 1) beschreibt im wesentlichen die nahezu
unendliche mikro-quantitative Variation, die in diesen Phänotypen meist nur noch schwer bzw.
nur noch mit besonderen Nachweismethoden zu erfassen ist. Dieser Teil der Variation spielt
erfahrungsgemäß weder in der
Züchtungsgenetik noch in der Makroevolution eine wesentliche Rolle. Das Gesetz der rekurrenten Variation
beschreibt somit die qualitative Variation sowie das Ausmaß der unmittelbar feststellbaren quantitativen
Variation. Die "Unendlichkeit im kleinen" wird zwar zum Teil in den fortlaufenden Mutationsexperimenten
ebenfalls beschrieben, sie ist jedoch für die Hauptfragen der Evolutionstheorie bedeutungslos, da sie
noch weniger als die qualitativen Veränderungen geeignet ist, über die primären Artgrenzen
hinauszuführen.
Trotz dieser und zahlreicher weiterer Einwände und
Tatsachen [vgl. die Links] missversteht die Synthetische
Evolutionstheorie (Neodarwinismus) die durch "Mikromutationen" (Mutationen mit
"slight or
even invisible effects on the phenotype" - Mayr) bedingte und jeweils auf Grenzen zulaufende Unendlichkeit
im
kleinen bis auf den heutigen Tag als
"ultimate
source" der
Makroevolution.
h) Prognosen und Retrognosen auf Grund des Gesetzes der rekurrenten
Variation
Die Rekombination mutierter Gene, die zu vorzeitig abgebrochenen Genwirkketten führen, sowie von Verlustmutationen in Regulator- und Target-Genen, verstärkt in der Regel die negativen Effekte auf die betroffenen homozygoten Individuen (Rekombinanten). Die Rekombination von neutralen bis schwach nachteiligen Allelen hingegen führt zur nahezu unendlichen Variation innerhalb der Art (wie wir das zum Beispiel von den Milliarden unterschiedlicher Phänotypen des Menschen her kennen). Und das ist auch von den Allelfunktionen her zu erwarten: "…we see in multiple alleles, that when genes mutate they generally continue to affect the same character in similar ways, in other words even genes are limited in their capacity for viable change" (Boyden).26b(2) Bei rekombinierten Genfunktionsverlusten addieren sich die Verluste, oder heben sich im Falle der selteneren Suppressormutationen zum Teil phänotypisch auf, - genotypisch addieren sich jedoch die Informationsverluste auch in diesem Falle weiter. Im neutralen Breich variieren und rekombinieren die schon bekannten Merkmale ebenfalls in mehr oder weniger umfangreichen Grenzen. "Mutations are merely hereditary fluctuations around a medium position" (Grassé). Die Rekombination solcher Allele kann nicht über die primären Artgrenzen hinausführen (vgl. auch Goldschmidts und Gottschalks Ausführungen oben).
Das gesamte Erfahrungsmaterial aus der Genetik und Züchtungsforschung ist mit diesen Aussagen in Übereinstimmung (Details siehe Artbegriff). Wir kennen keinen einzigen Fall primärer Artbildung durch Rekombination mutanter Allele, und die Rekombination mutativ entstandener neuer Gene entfällt ohnehin, da solche Gene nicht nachweisbar sind.
Zur Frage nach der Möglichkeit der Voraussage evolutiver Ereignisse schreibt Ernst Mayr: "Nichts in der Biologie ist vermutlich weniger vorhersehbar als der zukünftige Verlauf der Evolution." Und nach Aufführung von Beispielen, wie dem Werdegang der Reptilien des Perms, der Säugetiere und der kambrischen Fauna, fügt Mayr hinzu: "Unvoraussehbar ist auch die Evolution im kleinen Maßstab."26c
Als Hauptgrund vermerkt der Verfasser:
In biology, the pluralism of causations and solutions makes prediction probabilistic, if it is possible at all...the ability to predict is not a requirement for the validity of a biological theory .26d
Die Mendelschen Gesetze und das Gesetz der rekurrenten Variation bedeuten jedoch eine einschneidende Zäsur dieser Auffassungen: Die Möglichkeiten und Grenzen der Variabilität und der Evolution werden mit zunehmender Kenntnis der molekularen Grundlagen und der allelbedingten Phänotypabweichungen und Interaktionen im obigen Sinne prognostizierbar (thus, prediction is possible). Und niemand wird ernsthaft bestreiten, dass die Fähigkeit zur Voraussage ein bedeutender Gewinn für die Validität einer biologischen Theorie ist (vgl. die oben wiedergegebenen Definitionen zum Begriff GESETZ).
Für die Mendelschen Gesetze und das Gesetz der rekurrenten Variation probabilistisch bleibt jedoch die Antwort auf die Frage nach dem konkreten Fall innerhalb der Artgrenzen selbst in größeren Populationen: Welches mutante Allel, welche Rekombination wird sich - wenn überhaupt - in welcher Größenordnung in einer Population durchsetzen? (Das Hardy-Weinberg-Gesetz gilt nur für ganz bestimmte Bedingungen.) Und je kleiner die Population, um so geringer die Möglichkeit der konkret-individuellen Voraussage ("the pluralism of causations and solutions" - spielt in beiden Fällen sicher eine ganz erhebliche Rolle). Bei genauer Kenntnis genetischer Vorbedingungen und definitiver Umweltverhältnisse lassen sich jedoch auch in relativ kleinen Populationen zumindest die Möglichkeiten und Grenzen der Evolution bzw. der Degeneration vorausberechnen, und Voraussagen nach den Mendelschen Gesetzen (3 : 1 Spaltungen etc.) sollten möglich sein ("Vorausberechnung künftiger Ereignisse aus gegebenen Vorbedingungen").
Die prinzipielle Nicht-Vorhersehbarkeit der Makroevolution hingegen beruht grundsätzlich darauf, dass sie nicht auf Mutation und Selektion zurückzuführen ist.
i) Die Ursachen des Gesetzes der rekurrenten Variation
Die Gründe sind denkbar einfach. Es gibt nur eine begrenzte Zahl von Erbfaktoren, bei denen unter schrittweisem bis völligem Funktionsverlust (Allelbildung durch Mutationen, wobei wir die Gleichsinnkodons unberücksichtigt lassen) noch ein lebensfähiger, aber in vielen Fällen mehr oder weniger geschädigter Organismus gebildet werden kann. Stig Blixt hat dazu einige wertvolle Begriffsklärungen vorgenommen. Er stellt mit Ohno "verbotene Mutationen" in allen Genen fest, die für die Funktionen der Zelle und der Differenzierungsmechanismen eines Organismus essentiell sind und bei denen alle Mutanten mit potentiellen Phänotypabweichungen als völlig funktionsuntauglich sofort eliminiert werden (ein großer Teil der Mutationen in fast allen "house-keeping-genes" - Genen des allgemeinen Zellstoffwechsels - gehört dazu).27
Übrig bleibt also der variable oder redundante Teil der Gene, bei denen zufällige Sequenzänderungen abgewandelte, aber noch lebens- und konkurrenzfähige Phänotypen hervorrufen, und das ist aller Erfahrung nach nur ein kleiner Teil des Gesamtgenoms. Dieser Teil wirkt sich somit im neutralen bis schwach nachteiligen Funktionsbereich der Organismen aus. Er kann - in Übereinstimmung mit aller Erfahrung - jedoch keine völlig neuen anatomischen und/oder physiologischen Funktionen zwecks "Weiterentwicklung" durch Mutationen erschaffen. Der mutative Aufbau völlig neuer DNA-Sequenzen mit neuen Genfunktionen zur Durchbrechung des Gesetzes der rekurrenten Variation mit anschließender Artbildung ist bisher nirgends beobachtet worden und statistisch auch nicht zu erwarten. 28a
"To understand these observations one must clearly distinguish between two levels: 1) the level of the phenotypes and 2) the DNA level. On the latter, the potential of missense and nonsense mutations and other sequence deviations is nearly infinite. However, the spectrum of the resulting different phenotypes is not, because the space of functionally valid sequences within a given system of tightly matching regulatory and target genes and correspondingly co-ordinated functions involved in the formation of the finely balanced whole of an organism, cannot infinitely be stretched by chance mutations. To take a crude illustration: Drop your computer from the desk or take a screwdriver and a hammer, open the casing, shut your eyes and then forcefully operate in the innards! Depending on the number of computers and how often and for how long one proceeds to act this way, one may collect a nearly endless number of non-functional changes. Yet - with much luck - one may also select a few operationally diminished, but nevertheless still working, systems. Thus, one may demolish a computer in a thousand and more different ways by some accidental procedures. However, the resulting still more or less functional states (the functional phenotypes), will be limited. The hope to secure a Pentium III from a 486er by this method would be very bold indeed. - Of course, the situation in biology is more complex than in engineering, because organisms are, for instance, reactive entities. Nevertheless, limits to selection have repeatedly been found in several areas of biological research."28b
Die Evolutionstheoretiker A. Remane28c und A. Kühn28d haben zur Frage nach den Grenzen des Mutationsgeschehens schon vor mehreren Jahren einige nach wie vor voll zutreffende Bemerkungen vorgelegt, die die Problematik des Neodarwinismus weiter veranschaulichen (Hervorhebungen im folgenden Schriftbild wieder von mir); hören wir zunächst Remane:
Ein wichtiger Vorgang der Evolution ist die Bildung neuer Apparate, besonders bei Tieren. Das Auge der Wirbeltiere, die Leuchtorgane der Cephalopoden mit Linse, Tapetum und Reflectorschicht, auch viele Gelenke sind einfache Beispiele solcher Apparate. - In einem Apparat wirken verschiedene Teile verschiedener Herkunft koordiniert zusammen und erst die "passende" Gestaltung und koordinierte Leistung ermöglichen die Funktion des Organs. (Letzten Endes ist jeder Organismus ein einziger komplexer Apparat). Ein Gen und auch eine Mutation bedingen oft viele Merkmale und Leistungen (Polyphänie), aber diese sind nicht zu einem Funktionsapparat zusammengeschlossen. Ihre Gleichzeitigkeit ist durch die Leistungen des Gens bedingt, das in die Bildung vieler Merkmale eingreift. An der Bildung echter Apparate sind sicher ontogenetisch und phylogenetisch zahlreiche Gene beteiligt. Eine Ableitung aus unabhängigen Mutationen vom Typ der bekannten Mutationen stößt auf große Schwierigkeiten. Selbst wenn wir, wie bei Augen, eine successive Ableitung von einfachen Augenflecken morphologisch durchführen können, der Apparat also in aufeinander folgenden Schritten gebildet wurde, sind folgende Komplikationen notwendig: Der erste Schritt, die Anhäufung verschiedener Lichtsinneszellen an einer Stelle, ist leicht verständlich, der zweite Schritt, die Bildung von Pigment gerade an dieser Stelle, stellt an die nächste Mutation die Forderung einer strengen Lokalisation auf die Stelle des Augenflecks. Nur ganz bestimmte, seltene Mutationen könnten dieser Anforderung Genüge leisten. Das gilt für alle weiteren Schritte: Linsenbildung, Muskeln, Versenkung des Auges ins Innere usw., so daß ein enorm hoher Grad von Unwahrscheinlichkeit entsteht. Und ein solcher Vorgang hat sich nicht nur einmal ereignet; der Zufall kann ja im Einzelfall auch extrem Unwahrscheinliches zustande bringen (Ziehung des Großen Loses). In vielen Tierstämmen hat sich die Ausbildung solcher Apparate fast regelmäßig vollzogen. Kompliziert ist die Situation überall dort, wo Teile des Apparates ineinander greifen wie Räder in einem Uhrwerk oder wie Schloß und Schlüssel ineinander passen. Solche Strukturen gibt es bei Protozoen, Silicoflagellaten, Radiolarien und besonders bei Metazoen (Haftapparate, Chitingelenke u.a.) in Mengen. Die unabhängige mutative Abänderung eines der Teile stört die Funktion, und kooperativ wirkende Strukturmutationen kennen wir noch nicht.
Der ebenfalls abstammungstheoretisch
orientierte
Alfred Kühn
stimmt
Remane zu, wenn er schreibt:
Ob Häufigkeit, Richtung, Grad und Selektionswert der rein zufälligen Mutationen, die wir bis jetzt kennen, allein genügten, um die Gesamtheit der Formwandlungen der Stammesgeschichte zu bestreiten, läßt sich nicht beurteilen. Wir wissen nicht, ob die Bauplanunterschiede zwischen den höheren systematischen Kategorien, Ordnungen, Klassen, Tierstämmen, an deren allmählicher Herausbildung wir nicht zweifeln, auch durch die Summierung von Mutationsschritten, durch "additive Typogenese" zustande gekommen sind. Die Vererbungsforschung
bietet dafür keine Modelle. Die Neubildung komplizierter,
feinangepaßter Strukturen, wie z.B. Nesselkapseln, von
hochdifferenzierten Sinnesorganen (Linsenkameraaugen, Komplexaugen,
Gehörorganen), von elektrischen Organen, von wirksamen Zahnformen und
Zahnanordnungen mit entsprechenden Umänderungen an Muskulatur und
Schädelbau setzt das Zusammenwirken zahlreicher Gene voraus.
Alfred Kühn versucht dann diese Zweifel mit dem
Beispiel des
Pferdestammbaums zun relativieren - zu Unrecht, wie ich meine (vgl. zur
Unzulänglichkeit des Pferdestammbaums z.B.
die Ausführungen von
Junker und
Scherer 1998 und 2001, S. 232-235). Kühn fährt
dann jedoch fort: Die
Ausprägung
eines gesamten neuen
Leistungstypus, wie z.B. der Bienen-, Ameisen-, Termiten-arbeiterinnen,
einer Landform aus einer Wasserform, eines fliegenden Organismus,
erfordert eine Zusammenpassung (Koadaption) der verschiedensten, im Grunde
aller Organe des Körpers, einschließlich der nervösen
Grundlagen der Reflexe und Automatismen, unter denen es so raffinierte
Instinkthandlungen gibt wie die Orientierung und die
Verständigungsweise der Honogbienen. Solche harmonische Neu- und
Umkonstruktionen lassen sich durch Häufung und Naturauslese
gelegentlich auftretender Mutationen, wie wir sie bisher kennen, noch
nicht befriedigend erklären. Diese Kommentare erscheinen mir auch deshalb besonders
aufschlussreich, weil sie aus der Feder ehrlicher
Evolutionstheoretiker
stammen, die viel lieber gesagt hätten, dass alle grundlegenden
Probleme der Evolution längst mit Mutation und Selektion gelöst
seien (ähnlich zurückhaltend sind z.B. A. Portmann, J. Eccles, K. von Frisch, S. Vogel und viele
andere). Wie oben bereits ausführlich referiert, hat sich die
Situation in der
Zwischenzeit keineswegs zugunsten der
Synthetischen Evolutionstheorie verändert. Im Gegenteil, auch die
aufwändigsten und umfangreichsten Mutageneseexperimente haben nicht
über die primären Artgrenzen hinausgeführt und
sämtliche
von Remane und Kühn zitierten Beispiele liegen außerhalb
der durch Mutation erzeugten Variation, welche durch das
Gesetz der
rekurrenten Variation beschrieben wird. - Zur Diskussion weiterer
Beispiele vgl. H. Kahle: Evolution - Irrweg moderner
Naturwissenschaft? (4. Aufl. 2000). Zur Frage, wie neodarwinistische
Erklährungsversuche an synorganisierten Systemen gescheitert
sind,
vgl. Utricularia ,
die Diskussion zu Utricularia (sowie die daran
anschließenden
Kapitel 3 bis 5) und die Diskussionen von Michael
J. Behe.
Zum Redundanzthema zeichnet sich in den
letzten Jahren zunehmend noch eine ganz andere, wenn auch zu den
bisherigen Erkenntnissen komplementäre Möglichkeit zur
genetischen Erklärung der Grenzen der Evolution und des
mutationsgenetisch manchmal aufwändigeren Teils des Gesetzes
der rekurrenten Variation ab. Im
Rahmen meiner Infloreszenzstudien habe ich diese Frage wie folgt umrissen
(1999): Contrasting the spectrum of mutant
candidates which has appeared here with inflorescence structures which
have (for some cases, so far?) never appeared in all the Antirrhinum mutagenesis experiments of the 20th century taken together, it is to
be noted that all the functionally valid changes occur within the bounds
of raceme-like structures: No real (that is functionally efficient and
architecturally well-grounded) spadix-, catkin-, umbel-, capitulum-,
panicle-, thyrse/cyme-, mono- and/or polychasium-like configurations have
ever appeared.
It may be argued that the question, whether mutations
in or rearrangements of extant genetic material is sufficient or whether
functionally new special DNA sequences are necessary for the origin of
most of the very specific inflorescence structures just mentioned, cannot
be directly addressed in Antirrhinum or in Arabidopsis,
since neither of these species have ever displayed these features in the
wake of mutagenesis experiments. However, this 'negative evidence' is a
significant point for both, the practical as well as the theoretical
questions mentioned in the general introduction. First, it hints at
the limits of mutagenesis due (a) to possible redundancy
[of genes] or (b) absence of genes (relevant for the
question of the origin of the different inflorescence types). And, second,
in order to tag and clone the genes involved in the formation of such
inflorescence architectures and perhaps useful for man's optimal 'fruit
design', it is advisable to work with species already displaying these
features and possessing the corresponding gene functions (see future
developments). Es können also auch mehrere
weitgehend redundante Gene an der Ausbildung ein und desselben Merkmals
beteiligt sein (das Merkmal wird dadurch gut gegen Mutationen
"gepuffert"). Evolutionär liegt hier ein (weiteres) schweres Problem
vor, das Michael Denton28e in seiner Arbeit Nature's Destiny
(1998, pp. 337-
339) wie folgt umrissen hat (Hervorhebungen von mir): The strategy of using several different
means to achieve the goal is used in all manner of situations to guarantee
that the goal will always be achieved, even if one ore more of the means
fail. - Worauf instruktive Beispiele
für redundante Systeme, ja sogar mehrfach redundante Systeme, aus der
Raketentechnik und des Space Shuttles folgen. Zur genetischen Situation
bemerkt der Autor: It now appears that a considerable number
of genes, perhaps even the majority in higher organisms, are completely or
at least partially redundant. One of the major pieces of evidence that
this is the case has come from so-called gene knockout experiments, where
a gene is effectively disabled in some way using genetic-engineering
techniques so that it cannot play its normal role in the organism's
biology. A classic example of this came when a gene coding for a large
complex protein known as Tenascin-C, which occurs in the extracellular
matrix of all vertebrates, was knocked out in mice, without any obvious
effect. As the author of a paper commenting on this surprising result
cautions: "It would be premature to conclude that [the protein] has no
important function...[as] it is conserved in every vertebrate species,
which argues strongly for a fundamental role". The protein product of the
Zeste gene in the fruit fly Drosophila, which is a
component of certain multiprotein complexes involved in transcribing
regions of the DNA, can also be knocked out without any obvious effect on
the processes in which it is known to function.
The phenomenon of redundant genes is so widespread
that it is already acknowledged to pose something of an evolutionary
conundrum [Rätsel]. Although in the words of the author of one recent
article, "true redundancy ought to be, in an evolutionary sense,
impossible or at least unlikely", partially redundant genes are
common. As another authority comments in a recent review
article: Arguments over whether there can be true redundancy are moot for
the experimentalist. The question is how the functions for partially
redundant genes can be discovered given that partial redundancy is the
rule. ...Now, this phenomenon poses an additional challenge
to the idea that organisms can be radically transformed as a result of a
succession of small independent changes, as Darwinian theory supposes. For
it means that if an advantageous change is to occur, in an organ system
such as the nematode vulva, which is specified in two completely different
ways, then this will of necessity require simultaneous changes in both
blueprints. In other words, the greater the degree of redundancy, the
greater the need for simultaneous mutation to effect evolutionary change
and the more difficult it is to believe that evolutionary change could
have been engineered without intelligent direction. Redundancy also
increases the difficulty of genetic engineering, as it means that the
compensatory changes that must inevitably accompany any desired change
must necessarily be increased. Wir finden Redundanz auf allen
Ebenen: Erstens auf der DNA-Ebene (20% Gleichsinnkodons). Zweitens auf der
Proteinebene: Konservative Substitutionen durch Aminosäurereste mit
ähnlicher Funktion wie die ursprünglichen Aminosäuren (so
dass in einer Reihe von Fällen kaum oder keine Auswirkungen auf den
Phänotyp nachweisbar ist). Drittens: Darüber hinaus auf der
Genebene durch die soeben diskutierten redundanten Genfunktionen. Viertens
gibt es auch redundante Entwicklungswege (ontogenetisch) (vgl. Denton,
1998). Die Organismen sind also mit einer Vielzahl an
Systemen zum Schutz vor Mutationen ausgerüstet, wozu
(fünftens) die DNA-Reparatursysteme zu rechnen sind. Ein
Evolutionstheoretiker wie E.J. Klekowski Jr.28f, bemerkt zum
Thema
"Significance of Mutation" unter anderem: Although it is almost axiomatic that
mutation is fundamental for evolutionary change in plants (or any
organism), it does not necessary follow that the only (or even most
important) aspect of mutation in plant biology is evolution. It is
possible to construct an argument stating that the most important aspect
of
mutation is the selection for developmental and genetic systems with
maximum homeostasis. Such systems best resist the disruptive effects of
mutation and, therefore, stasis is promoted rather than evolutionary
change. (Schriftbild von mir.) Klekowski versucht damit vor allem, die
beobachtete Konstanz der Formen selektionistisch zu erklären -
bis zu einem hohen Grade ist das das genaue Gegenteil aller
neodarwinistischen Erklärungungsversuche, die ja bekanntlich auf das
Ziel hinauslaufen, die Entstehung aller Unterschiede und
Arten (und höheren Kategorien) im Reich des Lebendigen
naturalistisch verständlich zu machen. Um nur einen Punkt aus den
ungeklärten Fragen des selektionstheoretischen Postulats Klekowskis
herauszugreifen: Wie ist zum Beispiel die Entstehung der gene
conversion (vgl. Evolution durch Genduplikationen?) damit
mechanistisch zu erklären? - Bisher liegen jedenfalls weder für das
neodarwinistische 'Allerklärungspostulat' noch für die
mutationsgenetische Konstanzhypothese experimentelle Beweise vor.
Wäre es angesichts der Redundanz nicht besser -
wird vielleicht der eine oder andere Leser fragen - , mit Mutanten
für weitere Mutagenese-Experimente zu arbeiten, um überhaupt bis
zum den mutierbaren Kern eines Organismus durchzudringen? Die Antwort lautet: Genau das hat man
jahrzehntelang mit Hochzucht-
und weiteren Linien durchgeführt. Denn alle diploiden
Hochzuchtlinien sind bereits
Vielfach(verlust-)mutanten! Und an großen Mutantenzahlen besteht
trotz aller (Rest-)Redundanz kein Mangel. Bei noch voll redundanten
Genfunktionen
stößt
man jedoch erst bei Doppel- und/oder Vielfachmutanten auf die
"disruptive effects of mutation" - und auch dafür haben wir
zahlreiche Beispiele. Die entscheidenden Phasen des Ursprungs der
Organismenwelt kann man jedoch auch bei Genredundanz nicht durch
Genfunktionszerstörung erklären. Im Gegenteil, wie die
Ausführungen Dentons zeigen, weist die Redundanz selbst auf weitere
naturwissenschaftlich tragfähige Einwände gegen die Synthetische
Evolutionstheorie hin (von dem grundsätzlichen Problem der
Neuentstehung von Genen bei "Absence of Genes" einmal ganz
abgesehen). j) Bestätigung des Gesetzes der rekurrenten
Variation durch
Vavilovs Gesetz der Parallelvariation Der russische Genetiker und Agrarwissenschaftler Nikolaj
I. Vavilov hat 1922 aufgrund vergleichbarer Variationen bei verschiedenen Arten und Gattungen des Pflanzen- und Tierreichs "das Gesetz der homologen Serien in der Variation" formuliert, das er im Ansatz bis auf Mivart 1871 zurückführt. Er hebt folgende Punkte hervor:
Ganz allgemein bemerkt man beim Vergleichen von Mutationen verschiedener Pflanzen und Tiere generelle Linien der Variationen, sogar bei entfernten Gruppen von Organismen. (...) Zwergwuchs, Riesenwuchs, Albinismus und Verschmelzungen kommen im gesamten Pflanzenreich wie auch im Tierreich vor.29
V. Haecker bemerkt zur Frage nach der Verallgemeinerung der Beobachtungen einige Jahre später:
Zunächst zeigt ein Vergleich der verschiedenartigen Fälle von Parallelvariationen, daß der Grad der ,,Ubiquität", d.h. die Weite des Verbreitungsgebietes einer Potenz, sehr verschieden sein kann: Sie kann universell (ubiquitär i. e. S.), d.h. in sämtlichen Hauptgruppen der Tiere und Pflanzen da und dort in Erscheinung treten (Albinismus, Zwergwuchs), sie kann ferner eine kreis-, klassen-, ordnungs-, familien- oder gattungsweise (generelle) Verbreitung haben.30
Zu den Parallelvariationen führt Haecker weiter aus:
Was speziell diejenigen virtuellen Potenzen anbelangt, die als parallele Variationen in Erscheinung treten, so habe ich früher das Bild gebraucht, daß die betreffenden Qualitäten des Keimplasmas seinem "spezifischen" Kern, dem eigentlichen Artplasma, mehr oberflächlich und locker angegliedert sein müssen. Sonst wäre es ja nicht zu verstehen, daß die betreffenden Variationen mit den verschiedensten Artbildern verbunden sein könnten.31
Darüber hinaus hebt er hervor, dass es sich bei den weiter verbreiteten Parallelvariationen in der Regel um den mutativen Verlust von Merkmalen oder um solche leicht degenerativen bis biologisch indifferenten Charakters mit geringer korrelativer Bindung handelt. Der entscheidende Punkt bei diesen Beobachtungen ist nun, dass die parallelen Serien der rekurrenten Mutationserscheinungen auch bei nahe verwandten Formen - etwa Erbse (Pisum) und Platterbse (Lathyrus) - nicht die Art- und Gattungsunterschiede verwischen (sonst müsste ja die eine Art oder Gattung in die andere mutieren können).
Die Erbsenmutante Lathyroides beispielsweise erinnert schwach an die Platterbse (Lathyrus), ist aber völlig steril und mutiert laufend zum Wildtyp zurück.32 Typische Merkmale, wie die für viele Lathyrus-Arten geflügelten Stengel, sind dabei unter den Erbsen-Mutanten nie zusammen mit Fertilität aufgetreten. Solche Merkmale gehen also über das rekurrente Spektrum fertiler Mutanten von Pisum hinaus.32a
Generell kann man aufgrund aller empirischen Befunde nur feststellen, dass der Aufbau neuer physiologischer und anatomischer Systeme, durch welche sich primäre Arten und höhere systematische Kategorien voneinander unterscheiden, grundsätzlich außerhalb der rekurrenten Mutationserscheinungen liegen. Die Fülle von mutationsbedingten Parallelvariationen bei systematisch verwandten Formen, die jedoch nie die primären Artgrenzen überschreiten und dadurch die eine Form in die andere überführen, verdeutlicht noch einmal die Tatsache, dass auch bei weniger ubiquitär auftretenden Parrallelvariationen "die betreffenden Qualitäten des Keimplasmas seinem "spezifischen" Kern, dem eigentlichen Artplasma, mehr oberflächlich und locker angegliedert sein müssen. Sonst wäre es ja nicht zu verstehen, dass die betreffenden Variationen mit den verschiedensten Artbildern verbunden sein können" (Haecker). Die Überschreitung der Artgrenzen liegen also außerhalb der Mutantenspektren.
k) Mutationen beim Menschen
Heutzutage rechnen wir mit über sechs Milliarden Menschen. Wenn (nach den neuesten Schätzungen zu Beginn des Jahres 2001) bei 30.000 bis 40.000* Genpaaren und einer Mutationsfrequenz 10-5 pro Gen und Generation33 nun jedes einzelne Gen pro Generation mehr als 100.000mal mutiert, dann darf man davon ausgehen, dass in solchen Fällen in jeder Generation annähernd einmal das Potential zur Bildung erblicher Phänotypabweichungen durch Punktmutationen der Art durchgespielt und zu einem beachtlichen Teil auch freigesetzt wird. Beim Menschen wirken sich im Gegensatz zu den bekannten Daten aus der Pflanzengenetik die mutierten Allele von fast zwei Dritteln der festgestellten Loci dominant aus und sind damit schon beim ersten Auftreten phänotypisch festzustellen. Dennoch entwickeln sich nirgends neue Spezies des Homo sapiens, und, wie schon festgestellt, ist das bei der bekannten
"Qualität" der Mutationen auch nicht zu erwarten. Den Gesamtkatalog aller bisher beim Menschen festgestellten Mutationserscheinungen bringt A.V. McKusick34 laufend auf den neuesten Stand und zur Zeit zählt man über 5000 rekurrent auftretende erbliche Abweichungen. F. Schmidt kommentiert dazu: "Fast alle davon sind als Krankheiten oder Missbildungen im medizinischen Sinne des Wortes anzusprechen." Und ein paar Seiten weiter fährt er fort:Gewiß fallen Mutationen, die schwere körperliche und geistige Schäden verursachen, mehr auf und stellen demnach eine gewisse Interessantheitsauslese dar, weil die betreffenden Patienten früher oder später den Weg zum Arzt finden. Die Tatsache aber, daß Hunderten von Genetikern, die im letzten halben Jahrhundert genetische Untersuchungen am Menschen durchführten, in keinem einzigen Falle der Nachweis einer überzeugenden vorteilhaften Mutation gelang, ist ein klarer Beweis dafür, wie unsicher das Fundament ist, auf dem der Neodarwinismus einen wahren Wolkenkratzer an Spekulationen errichtet hat.35
Die Zahl der rekurrent auftretenden "Mutantentypen" nähert sich beim Menschen ebenfalls einem Limit. Nur ist im Vergleich zu den Pflanzen diese Zahl wegen der viel komplexeren physiologischen und anatomischen Organisation (und entsprechender Komplexität auf genetischer Ebene) wesentlich größer. Obwohl die Forschung bei den Pongidae (Menschenaffen) nicht annähernd so weit fortgeschritten ist wie beim Menschen, finden wir beim Schimpansen, Gorilla und Orang-Utan doch eine ganze Anzahl von Parallelvariationen (so zum Beispiel bekannte Trisomien wie das Down-Syndrom und viele zunächst beim Menschen beschriebene Punktmutationen), die uns wieder verdeutlichen, dass die bekannten Mutationsspektren mit ihren fast durchweg negativen Auswirkungen auf die betroffenen Individuen nichts mit der Überbrückung der Art- und Gattungsunterschiede zu tun haben können.
Die Mutantenspektren erklären weder den Ursprung der Formen noch die vielfach postulierte zukünftige "Weiterentwicklung". Die Mutantenspektren führen nicht über die primären Artgrenzen hinaus, ebenso wenig die Mendelsche Rekombination. "Die Spezies sind konstant", schlussfolgerte schon in den 50er Jahren des 20. Jahrhunderts der Genetiker Heribert Nilsson.36 Die anfangs zitierte Behauptung Mullers: "Nach Jahrmilliarden blinder Mutationen, die gegen die nachgebenden Grenzen der Umwelt anstürmten, entstand aus den Mikroben der Mensch" ist ein evolutionistischer Wunschtraum, der im schärfsten Kontrast zu der mangelhaften Qualität der Mutationen und dem Gesetz der rekurrenten Variation steht.
Zur Frage nach der zukünftigen Evolution neuer Menschenarten aufgrund von Mutationen und Selektion sind auch zeitgenössische Humangenetiker nicht gerade optimistisch. F. Vogel und A.G. Motulsky bekennen sich zwar zur Evolutionstheorie ("Human evolution is not yet finished. Evolution of the human species is not confined to the past."), aber auf die Frage nach der zukünftigen menschlichen Spezies bemerken sie zusammenfassend: "… its overall genetic composition will be similar to that encountered at the present."37 Sie modifizieren ihre Aussage leicht mit dem Hinweis auf Tendenzen zum Verlust rassischer und ethnischer Differenzen (das genaue Gegenteil der Artbildung!) und möglicherweise zunehmenden genetischen Defekten, aber auch besseren Diagnose- und Beratungsmöglichkeiten etc.. Aber mit evolutionistischen Voraussagen etwa für völlig neue und sich fortwährend weiterentwickelnde Menschenspezies auf der Basis der bekannten Mutationen halten sich die Autoren zurück.
Halten wir also fest: Es fehlt der naturwissenschaftliche Beweis, der es erlauben würde, die von vielen Genetikern mit "Unfällen" verglichenen Mutationserscheinungen zum "einzigen und alleinigen Grund jeglicher Neuerung und Schöpfung in der belebten Natur" zu erklären. Hingegen gibt es zahlreiche naturwissenschaftliche Gründe gegen die herrschende neodarwinistische Auffassung. Es ist zwar richtig, dass die Mutationen fast die einzigen bisher bekannten erblichen Veränderungen darstellen. Aber das gesamte Erfahrungsmaterial der Mutationsgenetik zeigt uns, dass die Schlussfolgerung von den negativen bis neutralen Mutantenspektren auf den Aufbau aller genetischen, physiologischen und anatomischen Systeme, auf die Entstehung aller Lebensformen, einen völlig ungerechtfertigten qualitativen Sprung enthält.
Zur Rettung der Synthetischen Evolutionstheorie hört man häufig die Erklärung, dass ja in der Erbsubstanz einer jeden Art ein in Jahrmillionen durchexperimentiertes System von Informationsträgern vereinigt sei und dass daher nur noch eine geringe Wahrscheinlichkeit für einen verbesserten neuen Genzustand beziehungsweise eine überlegene Genkombination bestehe. Der Rettungsversuch übersieht jedoch, dass auch bei den Organismen, die nach gängiger geologischer Zeitrechnung vor 100 oder 200 Millionen Jahren gelebt haben, schon "unvorstellbar zahllose Änderungsmöglichkeiten" im Laufe der vorangegangenen 400 Millionen beziehungsweise 1,5 Milliarden Jahre probiert und verworfen worden waren, so dass die Evolution nicht erst heute zum Stillstand gekommen sein müsste.
l) Die generelle Bedeutung des Gesetzes der rekurrenten Variation
Aus den mutationsgenetischen Befunden ist zu schließen, dass sämtliche Folgetheorien - vom philosophischen Materialismus über die evolutionäre Erkenntnistheorie bis hin zur modernen Soziobiologie - auf Sand gebaut sind. So originell, interessant und durchdacht solche Theorien auch sein mögen, wenn das Fundament nicht existiert, hängen auch alle darauf aufgebauten Schlussfolgerungen in der Luft. Das Fundament sind letztlich die Mutationen, die das Leben in all seiner Komplexität und Vielfalt erst möglich gemacht haben sollen. Leisten jedoch die Mutationen nicht das, was ihnen zugeschrieben wird, so fällt das gesamte darauf aufgebaute Gebäude an Spekulationen und Theorien in sich zusammen. Mehr als das rekurrent auftretende Mutantenspektrum kann die Selektion nicht selektieren. Mit den begrenzten Mutantenspektren sind auch die Möglichkeiten der Selektion limitiert - eine Beobachtung, die tausendmal bestätigt worden ist.
Keine Selektion und keine der Abermillionen von induzierten Mutationen, ob in Rekombination oder nicht, hat jemals über die primären Artgrenzen hinweggeführt. Um diesen Punkt noch einmal zu unterstreichen: "Neue Arten sind experimentell weder durch die schrittweise Anhäufung von Genmutationen noch durch die Induzierung einzelner progressiver Mutationen hergestellt worden. Hier liegt für die experimentelle Evolutionsforschung ein weites faszinierendes Arbeitsgebiet", schreibt ein führender neodarwinistischer Genetiker.38 Und das bedeutet, dass die intensive und umfangreiche Mutationsforschung des 20. Jahrhunderts mit Zehntausenden von Einzeluntersuchungen an den verschiedensten Arten kein brauchbares Ergebnis für die Artbildungstheorien gebracht hat. Meines Wissens rechnet heute kein Biologe mehr damit, durch richtungslose Mutationen noch neue Arten erzeugen zu können. Die Mutationsgenetik hat statt dessen deutliche Grenzen aufgezeigt - das Gesetz der rekurrenten Variation und die Selektionslimits. Das materialistische Weltbild ist ein Glaubensbekenntnis nicht etwa aufgrund der naturwissenschaftlichen Befunde, sondern gegen diese.
m) Der Darwinismus als Hindernis in der biologischen Forschung zur Zeit Mendels und heute
Als Mendel 1865 und 1866 seine Arbeit über die Vererbungsgesetze vorlegte, hatten seine Zeitgenossen kein Interesse an seinen Befunden. Ein Hauptgrund für die 35jährige Vernachlässigung von Mendels Befunden wird von M. W. Strickberger wie folgt beschrieben:
Nach Mendel änderten sich die betrachteten Faktoren während des Zeitraums der Beobachtung nicht, sondern drückten sich nur in neuen und unterschiedlichen Kombinationen in der Nachkommenschaft aus. Für diejenigen Biologen, die nach einer Ursache für die Variabilität in der Evolution suchten, bedeuteten Mendels Befunde das Gegenteil, nämlich eine unannehmbare Konstanz der Erbfaktoren. Hinzu kam, daß die Merkmale, die Mendel bei seinen Experimenten benutzte, Beispiele für diskontinuierliche Merkmale waren. Das heißt, Mendel beobachtete Unterschiede, für die es nur "alles oder nichts" gab, zum Beispiel lang oder kurz, glatt oder runzelig, gelb oder grün, usw. Viele Biologen, wie Darwin und andere, die sich mit Problemen der Vererbung befassten, suchten nach allmählichen Änderungen von einem Typ zum anderen, nach kontinuierlicher Veränderung.39
Außerdem war das Rekombinationspotential der konstanten Erbeinheiten nach den Mendelschen Gesetzen zwar groß, aber begrenzt und damit für die unbegrenzte Evolution unbrauchbar (beinhaltet also in jeder Hinsicht eine "unannehmbare Konstanz").
So standen die Mendelschen Gesetze nach Auffassung der meisten Biologen des 19. Jahrhunderts im scharfen Gegensatz zu den Voraussetzungen und Zielen der darwinistischen Evolutionstheorie und wurden vor allem deswegen nicht ernst genommen. Zum gleichen Ergebnis in dieser Frage sind viele weitere Autoren gekommen (de Vries, Bateson, Johannsen, Baur, R. von Wettstein, Kappert, Stern, Stubbe, Krumbiegel, Tinkle, Weiling, Jahn und andere; vgl.
mendel01.htm).Der erbitterte Widerstand der darwinistischen Schulen gegen die Mendelschen Gesetze zu Beginn des 20. Jahrhunderts - das ging soweit, dass die große britische naturwissenschaftliche Wochenzeitschrift Nature vorübergehend keine "mendelistischen" Befunde mehr zur Publikation annahm - bestätigt die Richtigkeit dieser Auffassung. Heute finden wir eine ganz ähnliche Situation. Die herrschende Theorie hat heute sowenig Interesse am Gesetz der rekurrenten Variation wie die Zeit Mendels an den später nach ihm benannten Vererbungsgesetzen (die im Rahmen der reproduzierbaren Rekombination ebenfalls eine Form der rekurrenten Variationen darstellen). Warum das so ist, zeigt der nächste Abschnitt.
n) Einwände des Neodarwinismus zum Gesetz der rekurrenten Variation
Das Gesetz der rekurrenten Variation mit seinen begrenzten Mutantenspektren und den daraus folgenden Selektionslimits steht heute ebenfalls im scharfen Gegensatz zur Synthetischen Evolutionstheorie. Aus dogmatischen sowie weltanschaulich-evolutionistischen Gründen ist daher Desinteresse, Unverständnis und/oder Ablehnung seitens der herrschenden Evolutionstheorie prognostizierbar:
"Nicht-phylogenetische Anschauungen, die phylogenetischen Ergebnissen widersprechen, sind für die Phylogenetik unbrauchbar", hat der Tübinger Botaniker W. Zimmermann40 bekannt und damit die Geisteshaltung der Biologie in unserer Zeit gekennzeichnet. Er bezog den Satz zwar zunächst auf die "idealistische Morphologie". Wie das Beispiel der Mendelschen Gesetze, die anfängliche Ablehnung des Hardy-Weinberg-Gesetzes und der Neutralen Theorie sowie Beispiele aus der Embryologie (Widerlegung des "Biogenetischen Grundgesetzes") und Paläontologie (Stichwort Punktualismus) etc. jedoch zeigen, werden de facto auch andere naturwissenschaftliche Ergebnisse (zumindest vorerst) als "unbrauchbar" klassifiziert, die der neodarwinistischen Phylogenetik und Evolutionslehre widersprechen. Aufgrund dieser Daten kann man Zimmermanns oben zitierten Satz berechtigt wie folgt umformulieren: Nicht-phylogenetische, naturwissenschaftliche Ergebnisse, die phylogenetischen Anschauungen widersprechen, sind für die Phylogenetik unbrauchbar. Da nun heutzutage fast alles Evolution und Phylogenetik ist, dürfte das Gesetz der rekurrenten Variation von neodarwinistischer Seite wohl erst einmal mit dem Prädikat "unbrauchbar" versehen werden. Sehen wir uns dazu die Einwände näher an, die der Mikrobiologe Herr Prof. K. in öffentlicher Diskussion mit mir 1990 formuliert hat:
Experimente zeigen, daß spontane wie induzierte Mutationsraten (Mutationen eines Typs pro Individuum, zum Beispiel Zelle, pro Zeiteinheit, zum Beispiel Generation, beziehungsweise Mutagendosiseinheit) für verschiedene Gene sehr verschieden sind. Spontane Raten variieren zum Beispiel von 10 -4 bis 10-10. Dadurch sind in Mutantensammlungen Typen mit hoher Rate meist mit mehr Exemplaren vertreten, die mit niederer, mit nur einem oder gar keinem.41
Diese Mutationsraten sind zum großen Teil aufgrund von Phänotypabweichungen anhand unterschiedlicher Nachweismethoden auf anatomischer und/oder physiologischer Ebene festgestellt worden. Da Gene unterschiedliche Redundanzgrade aufzuweisen haben, ist zunächst einmal ein großer Teil der Unterschiede der Raten auf die Funktionsfähigkeit der mutierten Gene in Relation zur Funktion des Gesamtorganismus zurückzuführen. Wenn in einem Protein sämtliche Substitutionen von Aminosäureresten für den Gesamtorganismus tödlich sind, findet man hier niemals irgendwelche homozygoten Phänotypen bei mutierten Genen. Sind in einem weiteren Protein 1 bis 2 Prozent der Aminosäurereste in einem bestimmten Abschnitt durch bestimmte andere mit Auswirkungen auf den Phänotyp substituierbar, so findet man entsprechend nur dann einen neuen Phänotyp, wenn genau in den entsprechenden codierenden Regionen der DNA die mutativen Veränderungen aufgetreten sind. Sind 10 bis 20 Prozent austauschbar, so werden die mutierten Gene (bei gleicher Mutationsrate) um so häufiger einen veränderten Phänotyp aufweisen. Kann der Organismus unter Phänotypänderungen ganz auf das Gen verzichten, dann wird jede mutative Funktionsbeeinträchtigung in diesem DNA-Abschnitt eine entsprechend häufige Mutante hervorrufen. Die unterschiedlichen "Mutationsraten" beruhen zum Teil auf den unterschiedlichen Toleranzgraden in der Funktion von Gen und Organismus bei mutativer Abänderung. Wir müssen also ganz klar die beiden Ebenen auseinanderhalten: die Mutation auf der DNA-Ebene (die zufällige Veränderung der Basensequenz) und das Resultat, die (homozygote) Mutante auf der organismischen Ebene, der Phänotypebene. Bei gleicher Mutationsrate kann in dem einen Fall niemals eine Mutante auftreten und in einem anderen ein veränderter erblicher Phänotyp (Mutante) bei fast jeder Funktionsstörung. Die Mutationsrate kann dabei völlig gleich sein. Modifiziert wird die Mutationsrate jedoch durch die Länge der Gene (lange DNA-Sequenzen können als Einheiten häufiger mutieren als kurze), unterschiedliche Erkennungsregionen für Transposons, Funktionsfähigkeit der Reparatursysteme und andere Faktoren.
Für unser Gesetz der rekurrenten Variation sind folgende Punkte entscheidend: Alle Gene mutieren. Die Anzahl der Gene ist begrenzt. Nur ein sehr geringer Teil der Genmutationen führt zu Phänotypänderungen, die für Züchtungs- und Evolutionsfragen von Interesse sein könnten. Diese Phänotypänderungen treten regelmäßig auf. Sie führen jedoch nicht über bestimmte morphologische und physiologische Grenzen hinaus. Daraus folgen die immer wieder festgestellten Selektionslimits.
Fazit der Züchtungsforschung: Aufgabe der Mutations-Programme bei Erreichen der Grenzen. Fazit für die Evolutionsfrage: Die Formen sind innerhalb dieser Grenzen konstant. - Kommen wir weiter auf Herrn Prof. K.s Einwände zu sprechen:
Das "Gesetz der rekurrenten Variation" ist leider bei Lönnig schlecht bzw. unverständlich definiert. Wenn gemeint ist, daß in Experimenten mit wachsender Zahl von Individuen, die mit einer Mutagendosis behandelt und auf bestimmte Mutationen, zum Beispiel "sichtbare" geprüft wurden, die Zahl verschiedener Einschrittmutantentypen ein Maximum anstrebt und also immer weniger bisher noch nicht bekannte Typen auftreten, so ist das eine selbstverständliche Folge der begrenzten Zahl von Genen in einem Genom und damit der Zahl möglicher Arten von Phänotypänderungen durch je eine Mutation pro Individuum.42
Wir sind uns also einig, dass die Zahl der Mutantentypen begrenzt ist und dass die Typen rekurrent auftreten. - Somit ist das Gesetz klar und verständlich definiert. Prof. K. schreibt weiter:
Wieso dies der evolutiven "Erzeugung immer neuer, sich weiterentwickelnder Phänotypen durch Zufallsmutationen" widerspricht, ist nicht zu sehen.43
Der Neodarwinismus will mit Mutationen die angenommene Entwicklung vom Prokaryonten bis zum Homo sapiens erklären. Ist aber die Zahl der Gene und der durch Mutation induzierbaren und brauchbaren Phänotypänderungen begrenzt - ob bei der "Mikrobe", der Pflanze oder dem Menschen - so widerspricht das ganz eindeutig der Idee der evolutiven Erzeugung immer neuer sich weiterentwickelnder Phänotypen! K. meint hingegen:
Denn nach der Additionstheorie geschieht die Weiterevolution zu höherer Komplexität und damit höheren Taxa durch die wachsende Zahl von Mutationsschritten im ganzen Genom, auch in schon früher mutierten Genen.44
Genau mit dieser falschen Annahme ist man in der Pflanzenzüchtung und den Versuchen zur "künstlichen Evolution" gescheitert! Wenn die postulierte Evolution nur eine Summierung von Mutationsschritten in verschiedenen Genen wäre, so müsste es ein Leichtes sein, mit den Zeitraffern der künstlichen Mutagenese (l000fach erhöhte Mutationsraten, bei Mikroorganismen sogar bis 10.000.000fach und mehr), der gezielten Rekombination und intelligenten Selektion laufend "Weiterevolution zu höherer Komplexität und damit höheren Taxa" zu erzielen. Statt dessen stoßen wir auf rekurrent auftretende Mutantentypen und auf Selektionslimits! - Weiter zu den Ausführungen von Herrn Prof. K.:
Damit steigt die Zahl der DNA-Varianten und somit der Phänotypen immer weiter und erreicht eine Grenze erst, wenn die millionenstellige Zahl möglicher DNA-Varianten erschöpft würde, was aber im ganzen All nicht realisierbar ist.45a
K. vermengt hier DNA- und Phänotypebene. Erstens: "Ca. 20 Prozent der Basenaustauschmutationen bilden wegen des degenerierten Codes ein neues Gleichsinncodon und damit den Wildtyp." Das heißt, diese Mutationen bilden keinen abweichenden Phänotyp. Zweitens: Wie oben schon erwähnt, lassen zahlreiche Aminosäurerest-Substitutionen erst gar keine Ontogenese zu und haben entsprechend auch keinen Phänotyp ("forbidden mutations"). Drittens: 99,99 Prozent und mehr der Phänotypänderungen sind für die Selektion nachteilig oder unbrauchbar und entfallen damit ebenfalls für die "Weiterevolution zu höherer Komplexität und damit höherer Taxa." Der Rest von 0,01 Prozent und weniger tritt gleich den anderen rekurrent auf, schafft aller Erfahrung nach aber auch nichts prinzipiell Neues und erreicht früher oder später sein anatomisches und physiologisches Limit.
Dass auch durch Rekombination keine neue Arten entstehen, das wussten schon die großen klassischen Genetiker wie Bateson und Nilsson (vgl. ausführliche Diskussion des Themas in der Artgegriffsarbeit). Man kann die Organismenwelt mit ihren unendlich komplexen und synorganisierten Strukturen nicht mit einer Summierung von neutralen bis schwach nachteiligen (zufälligen Gen-) Veränderungen erschaffen. Auch die von Neutralisten und Neodarwinisten inzwischen gleichermaßen akzeptierte Annahme, dass bei sämtlichen lebenden Fossilien die molekularen Uhren oft über 100 und mehr Jahrmillionen ununterbrochen weitergelaufen sind und dadurch in Zehntausenden von Genen einer jeden Art zahlreiche Nukleotid- und Aminosäurerestsubstitutionen ohne nennenswerte Änderungen der Phänotypen erfolgten, widerspricht der Additionstheorie.
Zur rekurrenten Variation bei Resistenzen, Genduplikationen, Polyploidie, Transposons und weiteren genetischen Phänomenen vgl. weiter Lönnig 1993, Kunze et al. 1997, Lönnig und Saedler 1997, und zur Frage der natürlichen Selektion, Lönnig 2001 (Anm. 7).
o) Schlussbemerkungen zum Neodarwinismus
Die Synthetische Evolutionstheorie behauptet, dass die Mutationen die Grundlage für jegliche Evolution durch Selektion seien. "Nach Jahrmilliarden blinder Mutationen, die gegen die nachgebenden Grenzen der Umwelt anstürmten, entstand aus den Mikroben der Mensch" (Muller). Nach dieser Theorie sind alle Organismen ohne Plan und Ziel, ohne Geist und Gott entstanden. Der Neodarwinismus ist die biologische Grundlage des Materialismus. Aufgrund einer naturwissenschaftlich-gründlichen Prüfung der behaupteten Leistungsfähigkeit der Mutationen sind zahlreiche Biologen und andere Naturwissenschaftler zu dem Ergebnis gekommen, dass die neodarwinistischen Behauptungen unbewiesen sind. Grundlegende Prognosen auf neodarwinistischer Basis haben sich in der Tier- und Pflanzenzüchtung, Embryologie und Paläontologie etc. als Fehlschlag erwiesen. Der Defektcharakter der meisten Mutationen sowie die Ableitung des Gesetzes der rekurrenten Variation und das Fehlen von Differenzierungsmutationen zeigen eindeutig, dass es sich beim Neodarwinismus nur um ein fragwürdiges Glaubensbekenntnis unter dem Deckmantel materialistisch missverstandener Naturwissenschaft handelt. Im Falle einer Konfliktsituation zwischen Neodarwinismus und biologischen Tatsachen und Gesetzmäßigkeiten haben Vertreter des Darwinismus und der Synthetischen Evolutionstheorie wiederholt versucht, ihr Glaubensbekenntnis in der Öffentlichkeit durch Abwertung und Unterdrückung der biologischen Tatsachen aufrechtzuerhalten. (Musterbeispiele: Mendelsche Gesetze, Hardy-Weinberg-Gesetz, Neutralitätstheorie vgl. mendel07.htm und Folgekapitel). Erst wenn sich nach langjährigen Anstrengungen die neu erkannten Gesetzmäßigkeiten nicht länger verschweigen und in die Synthetische Evolutionstheorie integrieren ließen, war man bereit, solche Tatsachen auch voll anzuerkennen.Nichts zeigt deutlicher den Ideologiecharakter des Darwinismus und der Synthetischen Evolutionstheorie als diese Tendenz, wissenschaftlichen Tatsachen und Gesetzmäßigkeiten grundsätzlich nicht die volle Bedeutung für konkrete wissenschaftliche Fragestellungen zuordnen zu können, solange solche Tatsachen nicht in das herrschende Glaubensbekenntnis integrierbar sind. Das Gesetz der rekurrenten Variationen dürfte dazugehören.
Auf jedem anderen Gebiet der Wissenschaft hätte man hingegen derart klaren und so häufig reproduzierten Befunden längst den Status einer allgemeinen Gesetzmäßigkeit zuerkannt.
Wenn aber Mutation und Selektion tatsächlich bei allen Lebensformen auf Grenzen stoßen, dann ist das materialistische Weltbild entweder falsch oder zumindest naturwissenschaftlich nicht abgesichert. Nur weil eine solche Schlussfolgerung für die meisten Zeitgenossen aus weltanschaulichen Gründen nicht akzeptabel ist, muss es sich bei den oben zitierten Befunden um vorläufige Ergebnisse handeln, denen man entsprechend wenig Bedeutung beimisst.
Falls Sie, lieber Leser, diese Einschätzung für überzogen halten, dann bitte ich Sie, meine - wie mir scheint - vergleichsweise freundlichen Ausführungen mit denen der nicht-(neo)darwinistischen Evolutionsbiologin Lynn Margulis von 1997 zu vergleichen (ich muss gestehen, dass selbst ich überrascht war, beim Studium des Buches derart starke Worte von einer überzeugten Evolutionsbiologin zu vernehmen, die jedoch die Symbiose für den Hauptfaktor der Enstehung der Lebensformen hält; - Hervorhebungen im Schriftbild von mir)45b:
More and more, like the monasteries of the Middle Ages, today's universities and professional societies guard their knowledge. Collusively, the university biology curriculum, the textbook publishers, the National Science Foundation review committees, the Graduate Record examiners, and the various microbiological, evolutionary, and zoological societies map out domains of the known and knowable; they distinguish required from forbidden knowledge, subtly punishing the trespassers with rejection and oblivion; they award the faithful liturgists by granting degrees and dispersing funds and fellowships. Universities and academies, well within the boundaries of given disciplines (biology in my case), determine who is permitted to know and just what it is that he or she may know. Biology, botany, zoology, biochemistry, and microbiology departments within U.S. universities determine access to knowledge about life, dispensing it at high prices in peculiar parcels called credit hours.
Und ein paar Seiten weiter (nach kritischen Hinweisen und Diskussionen zu "lessons in neo-Darwinian religious dogma" (S. 266), "the neo-Darwinian religious movement" (S. 270), "life, according to the neo-Darwinist gospel" (S. 272) und zu den Schlüsselbegriffen der Synthetischen Theorie - alphabetisch von 'Adaptation' bis 'Species', dass sie einem "quasi-religious purpose" dienen ("they bind practicing biologists into Fleckian thought-collectives that protect sacred knowledge", - S. 275) (und Recht hat sie in vielen Punkten!) fährt Margulis - unter Entwicklung ihrer "autopoietischen Sicht" - fort:
Yet the Academy guards, using neo-Darwinism as an inquisitory tool, superimpose a gigantic super-structure of mechanism and hierarchy that protects the throbbing biosphere from being directly sensed by these new scientists—people most in need of sensing it. The dispensers of the funds for scientific research and education and other opportunity makers, herd the best minds and bodies into sterile laboratories and white-walled university cloisters to be catechized with dogmatic nonsense to such an extent that many doctoral graduates in the biological sciences cannot distinguish a nucleic acid solution from a cell suspension, a sedimentary from an igneous rock, a kelp from a cyanobacterium, or rye from ergot.
In Europa gibt es dieselben Tendenzen (hin und wieder bis zum Fanatismus), - insgesamt jedoch scheint mir der Toleranzbereich hier etwas größer zu sein.
Noch stärkere Worte von der Verfasserin gegen die Synthetische Evolutionstheorie, sowie die Gegenkritik von führenden Neodarwinisten und weiteren Biologen, findet der daran interessierte Leser in dem Beitrag "Lynn Margulis: Science's unruly earth mother".45c
p) Alternativen
Was ist zu Ernst Mayrs Kommentar zu sagen, dass es keine brauchbare Alternative zur Synthetischen Evolutionstheorie gäbe? Er bemerkt:
It must be admitted, however, that it is a considerable strain on one's credulity to assume that finely balanced systems such as certain sense organs (the eye of vertebrates, or the bird's feather) could be improved by random mutations. This is even more true for some of the ecological chain relationships (the famous yucca moth case, and so forth). However, the objectors to random mutations have so far been unable to advance any alternative explanation that was supported by substantial evidence.46a
Dazu ist zunächst festzustellen, dass eine zweifelhafte und unwahrscheinliche Deutung nicht deswegen automatisch richtig wird, weil es im Moment keine gut abgesicherte Alternative dazu gibt. Wir kennen in den Naturwissenschaften zahlreiche offene Fragen, die eben solange offen bleiben müssen, bis im Zuge der weiteren Forschung eine voll befriedigende Erklärung gefunden wird.
Mit anderen Worten - und damit soll zugleich auf die Möglichkeit einer neuen Erklärungsdimension in der historischen Fragestellung zur Entstehung der Lebensformen aufmerksam gemacht werden: Soweit jemand auch bei der Ursprungsfrage als Erklärung in den Naturwissenschaften ausschließlich physikochemische Gesetzmäßigkeiten gelten lassen will, wird er sich angesichts der komplexen biologischen Tatsachen damit zufrieden geben müssen, dass die Hauptfragen zur Entstehung der Lebensformen noch offen sind.
Und mit den folgenden Sätzen wollen wir nun den direkten Sprung in diese soeben angedeutete, neue (und zugleich älteste) Erklärungsdimension der Menschheit zur Ursprungsfrage wagen und dabei die Frage nach ihrer logischen Berechtigung und Falsifizierbarkeit stellen:
Ist man jedoch bereit, sich vom evolutionistischen Denkverbot (frage auch bei den komplexesten und genialsten Konstruktionen in der Natur niemals nach dem Konstrukteur!) zu lösen, so könnte man folgende Argumentation akzeptieren. Der Zellphysiologe S. Strugger bemerkte einmal treffend: "Die Zelle ist das vollendetste kybernetische System auf der Erde. Alle Automation der menschlichen Technik ist gegen die Zelle nur ein primitives Beginnen des Menschen im Prinzip zu einer Biotechnik zu gelangen."46b Wenn nun schon "das primitive Beginnen" auf diesem Weg immer bewusstes Handeln, Intelligenz, Geist und Weisheit voraussetzt, - wie viel mehr muss das dann auf den Ursprung der tausendmal komplexeren kybernetischen Systeme der Lebensformen zutreffen!47 Sind die genial-komplexen kybernetischen Systeme der Organismen nicht bereits selbst "substantial evidence" für Intelligent Design? (Vgl. in diesem Zusammenhang auch die Arbeiten und Diskussionen von Michael J. Behe zum Begriff "irreducible complexity" nochmals:
http://www.discovery.org/crsc/fellows/MichaelBehe/index.html).46cDieses Denkmodell enthält im Gegensatz zum Neodarwinismus einen berechtigten "methodischen Sprung" - so berechtigt wie der Schluss vom Computer auf seinen Konstrukteur. Der Ansatz ist darüber hinaus grundsätzlich falsifizierbar:
Erstens: Durch den empirischen Nachweis, dass das Leben und die Information für komplexe Strukturen und Organe allein aufgrund physikochemischer Gesetzmäßigkeiten entsteht (Abiogenese und Bauplanbildung).
Zweitens: Durch den Nachweis, dass die kybernetischen Systeme der Organismen prinzipiell nicht mit den technischen vergleichbar sind, dass die Bionik auf einem falschen Ansatz beruht und dass für die Entstehung der Baupläne und höheren systematischen Kategorien kein koordinierter Informationsaufbau notwendig ist.
Drittens: Durch den Nachweis, dass die paläontologischen Befunde mit der neodarwinistischen Idee einer kontinuierlichen Entwicklung aller Lebensformen kongruent sind und dabei grundsätzlich einer diskontinuierlichen Entstehung von Familien, Ordnungen, Klassen und Bauplänen durch Intelligent Design widersprechen.
Viertens: Durch den Nachweis, dass alle komplexen Strukturen über Mutationen mit "leichten oder sogar unsichtbaren Wirkungen auf den Phänotyp", das heißt über Tausende von Zwischenstufen mit jeweils eindeutigen Selektionsvorteilen entstanden sind.
Bisher hat die Forschung in jedem Punkt das genaue Gegenteil aufgezeigt. Die Befunde sprechen ganz klar für Intelligent Design.
ANHANG
Wildarten, genetisches Potential und ParallelinduktionZur Frage nach Wildarten mit einem ursprünglich wesentlich größeren genetischen Potential folgen hier einige Auszüge aus der Artbegriffsarbeit von 1993, S. 473 und 586-588, und eine Ergänzung zu den Cichliden:
"Wir können damit feststellen, dass die umfangreichen Ergebnisse der Genetik, Pflanzen- und Tiergeographie, Paläontologie, Anatomie, Physiologie und Kybernetik uns ein übereinstimmendes Bild zum Artbegriff und zum Ursprung der Arten liefern. Alle Bereiche der Forschung zeigen uns, dass die Variabilität der primären Arten zwar ungeheuer reich, aber dennoch begrenzt ist. Darüber hinaus finden wir in Verbindung mit der Variabilität in der ganzen Organismenwelt Degenerationserscheinungen, die zur Bildung sekundärer Arten führen.
Mit dem Nachweis fortschreitenden
mutativen Abbaus
genetischen Potentials bei den Arten wird auch Licht auf das alte, bisher
jedoch völlig ungelöste genetische Problem der sogenannten
Parallelinduktion
geworfen.
"Parallelinduktion soll heißen,
daß parallel zum Soma auch
die Gene gleichsinnig abgeändert werden.…Wenn also in einem bestimmten Falle die Wirkung extremer Bedingungen gleichzeitig direkt die Reaktionskurve der Ausbildung eines Außencharakters beeinflußt und zugleich auch ein Gen in den Keimzellen quantitativ mutieren läßt, so wäre das Parallelinduktion. Das unverständliche dabei bleibt, daß gerade das Gen mutiert, dessen Reaktion auch modifikatorisch beschleunigt wird....Es ist wohlbekannt und seit den klassischen Arbeiten von NÄGELI viel diskutiert, daß Pflanzen, die in ein andersartiges Habitat gebracht werden, ihre Charaktere entsprechend ändern (Beispiel Ebenen- und Alpenformen); die Änderungen sind aber nicht erbliche Modifikationen. Andererseits gibt es in der gleichen Art Formen, die in verschiedenem Habitat erblich verschieden sind, und den für viele oder alle Formen des Habitats charakteristischen Typus zeigen, z.B. den Halophyten-Dünen-alpinen Typus. Die moderne Erklärung dafür ist, daß hier nicht etwa eine Vererbung erworbener Eigenschaften vorliegt, sondern eine Präadaptation. Das heißt, daß unter den zufälligen Mutanten der Stammform sich auch solche befinden, die im Gegensatz zu der Stammform die Bedingungen eines besonderen Habitats ertragen konnten und deshalb imstande waren, dort einzuwandern. In jüngster Zeit hat sich TURESSON ausführlich mit diesem Problem befaßt und kommt zu dem Schluß, daß diese Erklärung nicht ausreiche, sondern daß man annehmen müsse, daß die betreffenden Mutationen eine direkte Reaktion des Keimplasmas auf die spezifischen Außenbedingungen des Habitats
seien. Damit ständen wir wieder vor dem alten Rätsel der
Verursachung nützlicher Mutanten ohne Selektion" (Goldschmidt 1928,
p. 549). Was ist nun des Rätsels
Lösung? Stubbe wies im Rahmen seiner Antirrhinum-Studien
(1966, p. 154) darauf hin, "daß alle umweltbedingten Modifikationen
des Phänotyps auch mutativ bedingt sein können.'' Weiter
ließ sich zeigen, "daß zahlreiche, aber nicht alle mutativ
bewirkten Phäne auch als nicht erbliche Modifikationen, als
Phänokopien, auftreten können. Mit anderen Worten
läßt sich zeigen, daß alle modifikativen
Veränderungen, die durch Umweltverhältnisse verschiedener Art
bedingt sind, auch als Mutanten bekannt wurden,daß aber
nicht
alle Erscheinungsformen von Mutanten als umweltbedingte Modifikationen
phänotypisch wiederholt werden können" (Schriftbild
von mir). Goldschmidt ist zu den
gleichen Ergebnissen vor allem bei seinen Phänokopiestudien
(Drosophila) gekommen (1935 a und b, 1961). Die
ursprünglichen
Arten verfügten mit ihrem größeren genetischen Potential
über eine weite Anpassungsfähigkeit an alle möglichen
Umweltverhältnisse. Im Laufe der Zeit wurde diese weite
Anpassungsfähigkeit durch Anreicherung schwach nachteiliger Allele
(sowie Totalverlusten von am Standort redundanten Genfunktionen) an
den jeweiligen Arealen immer weiter eingeschränkt - mit
Ausnahme natürlich des für die spezielle Umweltbewältigung
notwendigen Teils. Andere Linien und Formen der selben Art sind jedoch noch nicht soweit degeneriert und verfügen entsprechend noch über eine größere Anpassungsfähigkeit. Durch den mutativen Abbau genetischen Potentials werden die Modifikationen mit der Zeit "erblich". - Das hat jedoch, so merkwürdig das zunächst vielleicht klingt, nichts mit der Vererbung erworbener Eigenschaften zu tun. Denn die Eigenschaften sind nicht evolutionistisch erworben worden, sondern waren von Anfang an mit der größeren Anpassungsfähigkeit gegeben. Aus diesem Anpassungspotential sind bei vielen Arten nur die für die jewei1igen Umweltbedingungen notwendigen Bereiche erhalten geblieben. Der 'Rest' ist durch Mutationen (Anhäufung schwach nachteiliger
Allele und sonstige Genfunktionsverluste) verloren gegangen - Bildung
sekundärer Arten. Durch
Rekombination noch vorhandener Wildtypallele (also von noch voll
funktionsfähigen Allelen) kann ein Teil des verlorengegangenen
Potentials zum Aufbau einer "neuen" Linie/Population
wiedergewonnen werden (vgl. p. 126 f.). Mutationen und Transposonaktivitäten steuern bei den sekundären Arten zur Variabilität und Mikroevolution bei." Nachträge
zur Parallelinduktion und Parallelvariation (ARTBEGRIFF,
S. 586-588)
"M.-W. Ho hat sich 1984 in dem Beitrag ENVIRONMENT AND HEREDITY IN DEVELOPMENT AND EVOLUTION (In: BEYOND NEO-DARWINISM. Eds. M.-W. Ho UND P.T. SAUNDERS; London) auf den Seiten 267-289 näher mit diesem Thema auseinandergesetzt. Wir lesen u.a.:
P.270: It turns out that parallels between artificially induced modifications and naturally occurring variations are extremely common. A very large body of observations ... show that structural divergences known or presumed to be hereditary are often correlated with environmental differences so much so that a number of ecological rules have been formulated on this basis. More importantly, however,these same structural modifications can be artificially induced in related organisms by simulating the appropiate environmental conditions in the laboratory.
(Hervorhebungen im Schriftbild von mir) - Worauf Beispiele aus dem Tierreich folgen. Schlussfolgerung:
In summary, the naturally existing variations - many of which are regarded as clear adaptations by Darwinists and neo-Darwinists alike - are adaptive to the very environment capable of eliciting the parallel artificial modification. This strongly suggests that the environment plays a central role in the origin and evolution of the adaptation itself.
P. 272: ... it is entirely plausible that adaptations should originate from interactions between organisms and environment [durch das gegebene Adaptations-Potential!]. The missing link is how the somatic changes resulting from those interactions could become hereditary in the sense that they seem to anticipate the environmental conditions for which they are an adaptation.
Da in der experimentellen Vererbungsforschung "alle modifikativen Veränderungen, die durch Umweltverhältnisse verschiedener Art bedingt sind, auch als Mutanten bekannt wurden" (vgl. Stubbe und Goldschmidt zitiert S. 473), lautet die Antwort auf diese Frage unter Berücksichtigung der Mutationsfrequenzen und dem Defektcharakter der meisten Mutationen (vgl. p. 340 ff): durch Einschränkung des ursprünglich größeren Anpassungspotentials! (Näheres p. 473.)
G. J. de Klerk nennt 1987 (Theor. Appl. Genet. 75, 223) bei der Besprechung von Matsudas Buch ANIMAL EVOLUTION IN CHANGING ENVIRONMENTS WITH SPECIAL REFERENCE TO ABNORMAL METAMORPHOSIS (New York 1987) zum Teil die gleichen Einwände wie Ho gegen die Idee der "genetischen Assimilation". Klerk schreibt:
... Matsuda's exclusive stress on environmental changes is most probably erroneous. Matsuda's second criticism of current evolutionary theory is that it does not recognize genetic assimilation. (The theory of genetic assimilation supposes that changes first occur at the phenotypic level and are then fixed in the genome by means of mutations which cause the same phenotypic effect.) ... it actually seems to be an illogical mechanism: why would organisms adapt at the level of the genotype, when they are already adapted at the level of the phenotype? If genetic assimilation does occur at all, its role is probably very limited.
Hos Einwand 1984, p. 272:
... as both the somatic modification and the coincident genetic mutation result in the same phenotype, how could natural selection distinguish between the two?
Alle drei Autoren irren sich hier: Die ganze von Goldschmidt oben beschriebene Problematik hat mit Evolution in neodarwinistischen Sinne wenig zu tun. Die genetische Assimilation beruht im wesentlichen auf der Degeneration (dem mutativen Abbau) nicht genutzten genetischen Potentials. Die Selektion spielt dabei nur insofern eine Rolle als sie die Funktionserhaltung des an einem Standort unbedingt notwendigen Teils des genet. Potentials garantiert.
Von Seiten der Molekularbiologie sind dazu in letzter Zeit bedeutende Beiträge zum Verständnis des Phänomens geliefert worden. So stellen U. Schibler und F. Sierra in ihrem Review 1987 ALTERNATIVE PROMOTERS IN DEVELOPMENTAL GENE EXPRESSION (Ann. Rev. Genet. 21, 237-257) die Frage nach der Funktion alternativer Promotoren und beantworten diese Frage u.a. wie folgt (p. 237):
In transcribing a single gene from multiple promoters an organism gains additional flexibility in the control of expression. In unicellular organisms these multiple promoters help to adapt the expression of a particular gene to the nutritional environment. In these cases, one promotor is often constitutive, whereas the other is inducible. (Beispiele...) ... In higher eukaryotes, alternative promoters are generally associated with tissue- and/or stage-specific genes. (Beispiele...) ... In several instances, the use of multiple promoters confers both qualitatively and quantitatively different expression of genes. Thus, in the parotid gland of both mouse and rat, amylase mRNA initiated at the upstream promoter of Amy-1 accumulates to a more than 100-fold higher concentration than mRNA initiated at the downstram promoter.
Bei der Erklärung des Phänomens kommen die Autoren nicht über evolutionistische Deutungen hinaus. Im Sinne der hier diskutierten Auffassung brauchen wir uns jedoch nur vorzustellen, dass die unter bestimmten ziemlich konstanten Umweltbedingungen lange Zeit nicht mehr gebrauchten Promotoren durch Mutationen funktionsunfähig werden und die Modifikation ist "erblich" geworden! Und es sei nochmals betont, dass das Phänomen nichts mit der Vererbung "erworbener Eigenschaften" zu tun hat. Stattdessen handelt es sich um die Degeneration eines ursprünglich größeren genetischen Potentials."
Cichlidae (Buntbarsche)
"All the incredible ecological, morphological and behavioural specializations that are found in these three groups of cichlids arose independently from one another. This would make these species flocks one of the most spectacular examples of convergence in all evolutionary biology" (Axel Meyer, Buchrezension, Nature 410, p.17; 1. März 2001; Hervorhebung im Schriftbild von mir).
Bei den Cichliden laufen mehrfach konvergente Artbildungsprozesse** in einem – geologisch gesprochen – so ungeheuer kurzen Zeitraum ab (Lake Victoria weniger als 14.000 Jahre, in mehreren Fällen sogar nur in wenigen Jahrhunderten), dass die mutative Neubildung von Genen, Genwirkketten und neuen Genfamilien als Differenzierungsursache gemäß allen bisherigen experimentalgenetischen und theoretischen Untersuchungen und Ergebnissen völlig auszuschließen ist.
"Da viele der heutigen Inseln, Schichtköpfe (kleinere felsige Erhebungen) und Uferbereiche mit ihren endemischen Fischfaunen bis vor wenigen Jahrhunderten also noch trockenes Land waren, dürfte die Etablierung eines Großteils der einzigartigen Buntbarschfauna des Malawisees in den letzten 200 Jahren stattgefunden haben" (G. Menting [2001]: Explosive Artbildung bei ostafrikanischen Buntbarschen. Naturwissenschaftliche Rundschau, August 2001, pp. 401-410; Zitat p. 404, Schriftbild von mir).
Wie aber sind dann die mehrfach konvergenten Differenzierungsprozesse in der Cichliden-Familie zu erklären?
Bevor wir auf diese Frage näher zu sprechen kommen, sollten wir uns noch einmal vergegenwärtigen, dass 1. die experimentelle Mutationsforschung – von Drosophila bis zur Mutationszüchtung – noch nie zu einer in der Natur beständigen neuen Art geführt hat, und 2. dass neben den Cichliden – in denselben Gewässern und demselben Zeitraum – zahlreiche andere Gattungen keinerlei neue Arten hervorgebracht haben. D.h. also: morphologisch praktisch völlig konstant gebliebene Formen existier(t)en neben den mehrfach unabhängigen Artenexplosionen der Cichliden ("they are more diverse than any other family of fish" - A. Meyer). Es wäre nun über den unzureichenden Zeitfaktor hinaus mutationsgenetisch überhaupt nicht nachvollziehbar, warum im Falle der Cichliden Mutationen durch funktionalen DNA-Aufbau ununterbrochen neue Arten und Gattungen produzieren sollten und dabei gleichzeitig die meisten anderen Tier- und Pflanzenformen, salopp gesprochen, völlig links liegen lassen.
Menting bemerkt zur Frage nach Erklärungsmodellen zum Ursprung der Mannigfaltigkeit innerhalb der Cichliden unter anderem (2001, p. 408):
"Neue Erklärungsmodelle explosiver Artbildungsprozesse basieren auf der Einschätzung, dass ein derart schneller und vielfältiger Wandel, wie er bei den Buntbarschen zu beobachten ist, für die Erzeugung neuer Gene oder Genwirkketten durch ein kleinschrittiges, akkumulatives Zusammenwirken von zufälligen Mutationsereignissen und natürliche Auslese keinen genügenden zeitlichen Spielraum lässt. Vielmehr wird angenommen, dass wir es bei der Radiation der Buntbarsche mit einem Prozess zu tun haben, der nicht allein durch äußere Faktoren (...), sondern maßgeblich auch durch innere Faktoren möglich wurde. Insbesondere ist hier an eine besondere genetische Ausstattung der Buntbarsche zu denken."
D.h. als Alternative zur mutativen Neubildung funktionaler DNA-Sequenzen bietet sich folgende Schlussfolgerung an: In derselben Zeit und demselben Raum verfüg(t)en die Cichliden über ein größeres genetisches Differenzierungspotential als andere Tier- und Pflanzenarten. Dieses Differenzierungspotential wurde im Laufe der Zeit wiederholt in die bekannte Fülle von morphologischen Arten und Gattungen umgesetzt. - Zu im Prinzip ähnlichen Aufassungen siehe G.R.Taylor 1983, J.C.Avise 1990, L.Seegers 1996, S. Scherer 1998, R.Junker und S. Scherer 1998 und 2001; und die Arbeitsgruppe A. Meyer beabsichtigt, die Frage experimentell zu untersuchen. - Hinweise und detaillierte Literaturangaben bei G. Menting 2001 (wie oben): Explosive Artbildung bei ostafrikanischen Buntbarschen. Naturwissenschaftliche Rundschau, August 2001. - (Siehe auch Lönnig 1986, pp. 186/187, 323/324, 402ff., 419, 473 [Hinweis auf die Cichliden Mexicos im Vergleich zu den ostafrikanischen Spezies und generell Bildung sekundärer Arten durch Abbau genetischen Potentials].)
Was heißt das aber für den vorliegenden Fall genetisch und molekularbiologisch? Genau an dieser Stelle stoßen wir auf die entscheidenden, aber zur Zeit noch offenen Fragen der Cichlidenforschung. Denn wir müssen bekennen, dass wir derzeit keine einzige DNA-Sequenz den funktional-morphologisch und ethologisch vielfach konvergent entstandenen Merkmalen der Cichlidenarten und –Gattungen zuordnen können.
Wenn wir einen Blick auf die Frage werfen, welche genetisch-molekularbiologischen Möglichkeiten für das postulierte größere genetische Differenzierungspotential derzeit zur Debatte stehen, so stoßen wir auf derzeit mindestens vier Möglichkeiten (vgl. weiter Lönnig 1993, Artbegriff [wie oben zum großen Teil schon zitiert] pp. 473, 545, 587/588).
Erstens: Das Vorhandensein alternativer Promotoren (je nach Bedarf werden Genfunktionen raumzeitlich unterschiedlich exprimiert).
Zweitens: Alternative Leserahmen (ein und dieselbe DNA-Sequenz kodiert mit Start- und Stopcodons an verschiedenen Stellen völlig unterschiedliche Gene).
Drittens: Gesteigerte Transposonaktivitäten (DNA-Transposons, die durch ihren „Cut-and-Paste"-Mechanismus Gene für alternative morphologische Merkmale und Verhaltensweisen an- und abschalten können) (Details zu Transposonfunktionen vgl. R. Kunze, H. Saedler und W.-E. Lönnig 1997: Plant Transposable Elements. Advances in Botanical Research 27, pp.331-470).
Viertens: Die ’normalen’ Mutationsprozesse schalten im Laufe der Zeit in den Cichliden-Populationen unterschiedlich funktionsfähige, aber für die Existenz der Gattung redundante Gene ab (also Abschalten durch die bekannten, häufigen Verlust-Mutationen). Das führte zu unterschiedlichen Differenzierungen, welche Tendenz noch durch die Rekombination mutierter Gene verstärkt werden konnte.
Alle vier Punkte sind in voller Übereinstimmung mit dem Gesetz der rekurrenten Variation und jeder einzelne Mechanismus würde zudem befriedigend das verblüffende Phänomen der Vielfachkonvergenzen erklären.
Auch wenn wir im Zuge der weiteren genetisch-molekularbiologischen Forschung die eine oder andere Möglichkeit beweisen können, so sind wir doch der Kernfrage, wie denn solche alternativ eingesetzten Promotoren, Lesenrahmen und Gene ursprünglich entstanden sind, noch keinen Schritt näher gekommen. Denn wie sollte die Evolution (gleichsam vorausschauend) mutativ zahlreiche alternative Genfunktionen entwickeln, die erst im Laufe der zukünftigen morphologischen (Art-)Differenzierung gebraucht werden?
Doolittle und Sapienza fassen die Problematik mit Hinblick auf die Frage nach der Funktion von Transposons wie folgt zusammen (und diese Zusammenfassung gilt auch für unsere Frage nach dem Ursprung eines größeren genetischen Potentials als zu einer bestimmten Zeit benötigt wird):
„Evolution is not anticipatory; structures do not evolve because they might later prove useful. The selective advantage represented by evolutionary adaptability seems far to remote to ensure the maintenance, let alone to direct the formation, of DNA sequences and/or enzymatic machinery involved."
Oder das Prinzip in den Worten von R. Dawkins: "In der Evolution hat bisher nichts anderes als der kurzfristige Nutzen gezählt; der langfristige Nutzen war nie wichtig. Es ist nie möglich gewesen, dass sich etwas entwickelt hat, wenn es dem unmittelbaren, kurzfristigen Wohl des einzelnen Lebewesens abträglich gewesen wäre" (worauf der Hinweis folgt, dass der Mensch insofern eine Sonderstellung einnimmt als er über diesen kurzfristigen Nutzen hinaussehen kann). Und man kann hinzufügen: Auch wenn wir begründet annehmen können, dass gewisse Nachteile sogar kurz- und langfristig möglich waren (Tolerieren von schwach nachteiligen Allelen, Anhäufung von junk DNA, Degeneration im Organismenreich etc. - vgl. Artbegriff pp. 403 ff.), so ist es jedenfalls nie möglich gewesen, dass die Evolution vorausschauend eine ganze Fülle komplexer genetischer Informationen, die kurzfristig völlig überflüssig gewesen wäre, für das langfristige Wohl und die zukünftige Bildung von Arten und Gattungen erzeugt hätte. Denn das hätte die Bildung reichhaltiger genetischer Information zunächst ohne jede Selektion und zudem kurzfristig ohne jede morphologisch-physiologische Funktionen bedeutet.
Wie aber soll dann die Information für die alternativen und ursprünglich nicht unmittelbar notwendigen (d.h. redundanten) Genfunktionen entstanden sein?
An dieser Stelle drängt sich dem einen oder anderen Leser vielleicht doch wieder der Gedanke nach finalen Ursachen auf. Da nun aber – wie oben schon zitiert - "teleologisch-finalistische Interpretationen bekanntlich naturwissenschaftlich nicht erlaubt sind", müssen sich die meisten zeitgenössischen Denker wohl derzeit mit damit begnügen, dass die Frage nach dem Ursprung des umfangreichen genetischen Alternativpotentials der Cichliden noch offen ist.
Jedenfalls steht das vielleicht spannendste Kapitel der Cichlidenforschung, die Klärung der genetisch-molekularen Grundlagen der vielfach konvergenten Artdifferenzierung sowie der genauen Ursachen der Bildung der originären DNA-Sequenzen samt raumzeitlicher Genexpression noch aus.
Was auch immer sich als genaue Ursache herausstellen wird - ob eine oder mehr der vier oben zitierten oder gar ganz neue Möglichkeit(en) zur Bildung eines umfangreichen (über die kurzfristigen Bedürfnisse der ursprünglichen Cichlidenpopulation erstaunlich weit hinausgehenden) genetischen Potentials - fest steht, dass die "Urformen" oder Ausgangslinien der Cichlidenfamilie tatsächlich aus "polyvalenten Stammformen" (Junker und Scherer 1998, 2001) bestanden, so dass sich die Frage nach teleologisch-finalen Interpretationen unausweichlich stellt.
Dr. G.s Kommentar zur Teleologie-Frage (Dr. G. ist Biologe):
Wenn nach L.-C. Schulz
teleologisch-finalistische Fragestellungen in der Biologie unzulässig
sind, ist die Aussage, dass der Mensch ein Zufallsprodukt sei,
natürlich auch unzulässig. Diese Behauptung beantwortet die
Sinnfrage eben doch, dass es nämlich keinen Sinn gäbe.
Evolutionisten haben die Sinnfrage nach dem Leben eben
schon vor ihrer Forschung beantwortet: Es gibt nur den Zufall
(1). Keineswegs lässt also der Evolutionist diese Frage offen. Es gibt genau diese vorweggenommene Deutung des Lebens
auf dieser Erde, die diesen Forschern die objektive Erkenntnis und
Beurteilung des Bios auf dieser Erde verwehrt. Die Frage nach dem Leben auf dieser Erde ist für
Evolutionisten primär eben keine biologische, sondern eine
weltanschauliche Frage. Ihr "Trick" ist es, diese eher philosophisch
bzw. theologische Frage nicht mit den erkenntnistheoretischen Methoden
dieser Disziplinen zu lösen, sondern sie durch Fakten der Biologie
(Paläontologie, Botanik, Zoologie, Anthropologie, Genetik,
Molekularbiologie) zu belegen (2). Da diese Fakten aber ihre Theorie nicht belegen
können, werden die Fakten gedeutet in immer wieder neuen
Evolutionstheorien, die natürlich alle ihre Fehler haben, weswegen
bald eine neue Theorie herhalten muss. Die einander ablösenden
Theorien werden als Fortschreiten der Wissenschaft bezeichnet, aber in
Wirklichkeit ist es nur ein Durchspielen von Fehlerbehebung unter
Einbeziehung neuer Fehler, auf die die Vorgänger noch nicht gekommen
sind. (Dr. G.s Kommentar zur Aussage: "Der Mensch war bei
diesem Mutationsgeschehen weder geplant noch überhaupt in
irgendeiner Weise vorauszusehen...":) Unterstellt man eine reine
Zufallsentstehung von Lebewesen, gibt es keinen Grund anzunehmen, dass in
einer anderen Evolutionsgeschichte überhaupt Lebewesen entstehen und
wenn, warum dann Fauna und Flora - die Entstehung des Menschen wird ja
auch negiert. Muller (3) zeigt, wie er als Geschöpf nur in der
Ordnung einer Schöpfung denken kann. Eine reine Zufallsentwicklung
ist also ein für den Menschen undenkbarer Gegenstand.
____
Zu (1): (Ergänzungen von mir,
W.-E.L.): "...und
die
Naturgesetze." (2) Ich würde sagen: "Ihr "Trick" ist es, diese
vorweggenommene Deutung der Entstehung des Lebens auf dieser Erde als die
einzig mögliche naturwissenschaftliche Erklärung hinzustellen
and dann
fälschlich zu behaupten, dass nur diese den Fakten
entspricht" etc.. - Die totalitär-materialistische Deutung des
Lebens
ist natürlich auch eine philosophische, ja sogar eine theologische
Aussage über das Wesen der Dinge (theos: Gott; logos: Lehre,
Wort). Die naturalistische Theologie (Gotteslehre) ist jedoch samt ihrer
dogmatisch-erkenntnistheoretischen Methodik ("teleologisch-finalistische
Fragestellungen sind in der Biologie
prinzipiell unzulässig") nachweislich
unzureichend. Das macht selbstverständlich die
herkömmlich-konservative Theologie - gegen deren Methodik und
Dogmatik
ich wiederum viele
Bedenken anmelde - nicht automatisch richtig. Aber wir müssen uns
nach
einer über den Naturalismus hinausgehenden Alternative umsehen. Zu
(3): ...und viele andere zeigen. Dr. H. (ebenfalls Biologe): Was ist
über die
Geschichte des Gesetzes
der
rekurrenten
Variation zu berichten? W.-E.L.: Der Gedanke, dass die Ergebnisse
der
Mutationsforschung
in einer
phänotypischen Gesetzmäßigkeit zusammengefasst werden
könnten, ist mir das erste Mal bei der Auswertung von mehreren
Tausend Pisum-Pflanzen auf dem Versuchsfeld des Genetischen
Instituts der
Universität Bonn auf dem Venusberg im Sommer 1982 gekommen: Ein
wesentlicher Anstoß bestand in dem Phänomen, dass vier
unabhängig voneinander entstandene Mutanten, die in mehreren Genen
mutiert waren, nach Kreuzung mit der Kulturvarietät "Dippes gelbe
Viktoria" (Ausgangslinie für drei der Mutanten) immer wieder ein ganz
erstaunlich ähnliches Spaltungsmuster innerhalb eines bestimmten
phänotypischen Spektrums zeigten (F2
bis F6
wurden untersucht); es
waren also mehrfach unabhängig voneinander dieselben oder
unterschiedliche Gene mit ähnlichen phänotypischen Auswirkungen
mutiert, und zwar immer mit Auswirkungen innerhalb ganz bestimmter
phänotypischer Grenzen. Die Pisum-Ergebnisse wurden 1985
publiziert
(Genetica 66: 29-40), die darin zum Ausdruck kommende allgemeine
genetische
Gesetzmäßigkeit jedoch noch nicht direkt angesprochen.
Ein
wohl weiterer Anstoß bestand im Kommentar eines
Pisum-Mutationsgenetikers, der seit Jahrzehnten Mutationen
bei der Gartenerbse induziert hatte, zu meinem Anliegen, erneut Mutationen
bei
Pisum
zu induzieren, dass man dabei praktisch nichts Neues mehr erwarten
könne (es würden nur wieder die gleichen Chlorophyll- und
Internodienmutanten etc. auftreten). Schließlich sei als dritter
Anstoß die Begrenztheit der Phänotypen des für mich damals
ersten direkter Beobachtung und genetischer Feldarbeit
zugänglichen
größeren
Mutantensortiments genannt (wiederum Pisum), sowie die Tatsache,
dass bei den zahlreichen Mutanten und Rekombinanten
von
Artbildung nirgends die Rede sein konnte (ja hier waren nicht einmal neue
Kulturvarietäten, die den Anerkennungskriterien des
Bundessortenamts entsprachen, durch die Vielzahl von Mutationen und
Rekombinationen erzeugt worden).*** Weiter zeigte mir das Studium der umfangreichen
Literatur zur
Mutationsgenetik, dass meine Beobachtungen und der oben genannte Kommentar
des erfahrenen Pisum-Genetikers keineswegs Ausnahmefälle
beschrieben. Vielmehr
war das Phänomen, dass die in mutationsgenetischen Experimenten
induzierten Phänotypen gegen ein Limit laufen, praktisch schon seit
Jahrzehnten in der Mutationsgenetik rund um den Globus immer wieder
festgestellt worden. Unter der ideologischen Vorherrschaft des
Neodarwinismus, der den Ursprung
der gesamten Lebensvielfalt mit Mutationen erklären möchte,
konnte sich
jedoch keiner der mit der Mutationsgenetik befassten evolutionistischen
Forscher zur Erkenntnis einer allgemeinen genetischen
Gesetzmäßigkeit mit dem Hauptergebnis limitierter
phänotypischer
Variation durchringen, denn das hätte die Aufgabe
der Synthetischen Evolutionstheorie bedeutet! Der Ansatz zur Erkenntnis
eines solchen genetischen Gesetzes findet sich jedoch implizit bei
antidarwinistischen Genetikern wie Nilsson und Lamprecht, er wird dort
aber nicht explizit formuliert und benannt. Von den älteren Züchtern
postulierte der lamarckistisch und 'antimendelistisch' eingestellte Luther
Burbank (1849-1926) (in der
amerikanischen Literatur als
"the greatest breeder"**** des 20. Jahrhunderts bekannt) sogar ein
Gesetz zur Limitierung der Variation. Er schrieb: "I know from my
experience that I can develop a plum half an inch long or one two and a
half inches long, with every possible length in between, but I am willing
to admit that it is hopeless to try to get a plum the size of a small pea,
or one as big as a grape-fruit....In short, there are limits to the
development possible, and these limits follow a law....It is the
law [of the reversion to the average]. Experiments carried on
extensively
have given us scientific proof of what we already guessed by
observation: namely, that plants and animals all tend to revert,
in successive generations, towards a given mean or average...In short,
there is undoubtedly a pull toward the mean which keeps all living
things
within more or less fixed limitations..." (Zitiert nach W.R.Bird: THE
ORIGIN OF SPECIES REVISITED; Vol I, 1989, p. 87; bold von
mir). Wenn wir davon ausgehen, dass "a pull toward the
mean" sich heute schon weitgehend kausalanalytisch
definieren lässt, dann waren diese Statements zwar als völlig
realistische
Beschreibung seiner (Burbanks) lebenslangen Studien und Beobachtungen zur
Variation ein
großer Schritt in die
richtige Richtung. Sie waren aber dennoch mehr eine allgemeine
Feststellung als ein
genetisch gefasstes und begründetes Gesetz. Seine Aussagen
hätte er allerdings mit
den Mendelschen Befunden bis zu einem gewissen Grade kausal fassen und
ausbauen können
(z.B. begrenzte Zahl von Genen, Presense-Absense-Theorie, Reversion
durch Mendelsche Rekombination). Da aber
damals von Transposons noch nichts bekannt war, ist diese
Anmerkung für die Beobachtung der Reversionen nur zum Teil
relevant. Das erste Mal habe ich die
mutationsgenetische
Regelhaftigkeit und
Begrenzheit der Phänotypen unter Aufführung zahlreicher Daten in
meiner Artbegriffsarbeit von 1986 als "Regel der rekurrenten
Variation" bezeichnet und publiziert. Im Jahre 1995 folgte dann nach
zahlreichen weiteren Beobachtungen und Bestätigungen sowohl durch die
eigene
(recht umfangreiche) empirische Arbeit mit Pisum und
Antirrhinum als auch
durch das fortgesetzte Studium der mutationsgenetischen Literatur die
Veröffentlichung unter dem Titel: MUTATIONEN: DAS GESETZ DER
REKURRENTEN VARIATION (pp. 149-165 in: Streitfall
Evolution. Hrsg.: J. Mey, R. Schmidt und
S. Zibulla. Universitas. Wissenschaftliche
Verlagsgesellschaft. Stuttgart). Die hier vorliegende, verbesserte und
stark erweiterte Internetversion erfolgte im Frühjahr 2001.
EINWAND: "Aber bedenken Sie doch die
Jahrmillionen, - da war die Artbildung über die Grenzen der
Mutationsspektren hinweg eben doch möglich."
Zu diesem Einwand bitte ich, wohlwollend
die folgenden klärenden Zitate und
Kommentare zur Kenntnis zu nehmen (auch wenn diese vielleicht etwas zu
direkt formuliert sind):
J.H.Fabre: "Wenn die Schwierigkeiten zu
groß werden, flüchtet man hinter die Nebel der Jahrhunderte
[und Jahrmillionen1], versteckt man sich, soweit die Phantasie
es
gestattet, im Dunkel der Vergangenheit, ruft man die Zeit an, diesen
Faktor, über den wir so wenig vermögen und der sich gerade
deshalb so gut dazu eignet, Phantastereien zu verhüllen."
(W.-E.L., Augenschrift, pp. 59/60): Wie
ungerechtfertigt die Verabsolutierung von Variation und Selektion ist,
darüber haben wir ja schon einiges gehört. Bleibt noch das Spiel
mit den Jahrmillionen, zu dem Andermann einmal nach gründlicher
Untersuchung festgestellt hat, dass es im Grunde genommen von "vornherein
eine faule Angelegenheit ist, wenn man zu "unendlich
langen" Zeiträumen und zu einer unkontrollierbaren Vergangenheit
Zuflucht nehmen muss. Wir sehen immer wieder denselben Versuch, logische
Schwierigkeiten dadurch abzuschwächen, indem man das Problem
verschiebt und sich auf die Wirkung der Zeit hinausredet."
M.Eden bemerkt zu demselben Punkt: "Die Länge der
Zeit ist nur dann sachdienlich, wenn die Wahrscheinlichkeitsstruktur der
Ereignisse und Veränderungen, die sich in dieser Zeit abspielen,
ebenfalls bekannt ist" (Mathematical Challenges to the Neo-Darwinian
Interpr. of Evolution; 1967, S. 8).
(W.-E.L., 1. Staatsexamensarbeit, pp. 84/85): An
diesem Punkte [mehrfach konvergente Evolution komplexer Strukturen]
hört man nun
häufig den Einwand, dass dieser
Prozess ja ungeheure Zeiträume, Jahrmillionen, voraussetzt, er also
experimentell nicht zu erfassen ist - Schindewolfs oben zitierter
Einwand. Auch der Neodarwinist Dobzhansky meint in dieser Hinsicht (1957,
p. 388):
"The applicability of the experimental
method to the study of such unique historical processes is severely
restricted before all else by the time intervals involved, which far
exceed the lifetime of any human experimenter. And yet it is just such
impossibility that is demanded by antievolutionists when they ask for
"proofs" of evolution which they would magnanimously accept as
satisfactory."
Man muss schon recht kritiklos sein, um
sich mit einer solchen Erklärung zufrieden zu geben und zu meinen,
dass damit alles Wesentliche erklärt sei. F.Andermann hat schon 1938
[Irrtum und Wahrheit in der Biologie] zu
dem Zeitproblem ein paar heute noch, wie ich meine, voll gültige
Anmerkungen gemacht, die den Kern der Sache so vollendet treffen, dass ich
ein ausführliches Zitat von ihm hier wiedergeben möchte. Er
schreibt (p. 242):
"Rein logisch spielt die Zeitfrage beim
Problem der
Artbildung überhaupt keine Rolle. Ob sie rasch oder langsam vor sich
geht, ist nebensächlich. Vorerst muß sie als Tatsache
überhaupt erwiesen sein. Würde uns die Erfahrung etwa zeigen,
dass sich ein Schaf im Laufe von zwei oder drei Generationen in ein
Nashorn umwandeln kann, so müßten wir das als Tatsache
hinnehmen, nicht weniger begreiflich wie jede andere Metamorphose in der
Natur (die wir in keinem Falle wirklich "verstehen"). In der individuellen
Entwicklung, von der allein es wirkliche Erfahrung gibt, verlaufen die
einzelnen Phasen der Metamorphose mit verschiedener Geschwindigkeit.
Die Schmetterlingslarve behält ihre Form lange
Zeit hindurch bei, ohne auffällige Veränderungen
durchzumachen. Tritt sie aber ins Puppenstadium, so verläuft die
weitere Umwandlung zur Imago sehr rasch, gleichsam über
Nacht. Wüßten wir nicht, daß der flatternde Schmetterling
mit den prächtigen Flügeln einmal ein häßlicher Wurm
war, und würde nicht diese Verwandlung vor unseren Augen vor sich
gehen, wir g1aubten bestimmt nicht daran und würden eine solche
Behauptung für ein Märchen halten.
Wie töricht ist doch der Mensch, der da glaubt, es
müsse in der Natur genau so hergehen, wie er es sich
vorstellt. Gerade das Beispiel von der Umwandlung der Larve in die Imago
zeigt uns sehr deutlich, daß die Natur keinesfalls den Weg der
allmählichen Evolution gehen muß, wie ihn die
Entwicklungstheoretiker ihr vorschreiben. Und in gewissem Sinne kann man
hier sagen, daß zwei Arten auseinander entstehen, ohne daß sie
die geringste Ähnlichkeit miteinander haben. Was zwingt uns also zur
Annahme, daß eine Art nur aus einer ihr ähnlichen entstehen
muß, und - was wissen wir überhaupt davon, wie Arten in der
Natur entstehen?" Und auf Seite 34 lesen wir: "Wenn
nun wirklich in der
Natur die Tendenz bestünde, die niederen Arten in die höheren zu
überführen, so hätte sie bis jetzt reichlich Gelegenheit
gehabt, es uns zu zeigen. Merkwürdig genug, daß sie es
während der ganzen geschichtlichen Beobachtungszeit nicht getan
hat. Und so ist es von vornherein eine faule Angelegenheit, wenn man zu
"unendlich langen" Zeiträumen und zu einer unkontrollierbaren
Vergangenheit Zuflucht nehmen muß. Wir sehen immer wieder denselben
Versuch, logische Schwierigkeiten dadurch abzuschwächen, indem man
das Problem verschiebt und sich auf die Wirkung der Zeit
hinausredet."
(W.-E.L., Augenschrift, p. 84): Wenn "die
Evolution" als Naturgesetzmäßigkeit so etwas [die Bildung
komplexer Strukturen wie die des Auges] durch Mutation und Selektion kann
(und das mehrmals zustande gebracht hat), dann sollte das auch prinzipiell
reproduzierbar sein! Das an solchen Stellen übliche "Hinausreden auf
die Wirkung der Zeit" ist übrigens ein völlig
haltlos-unwissenschaftliches Ausweichmanöver, solange man nicht die
Wahrscheinlichkeitsstruktur der Ereignisse und Veränderungen, die
sich in dieser Zeit abspielen, kennt (vgl. S. 59/60). Denn jeden Unfug
könnte man behaupten, - etwa die Entstehung griechischer Tempel durch
geologische Faktoren (ein Beispiel Wigands), die plan- und ziellose
Erzeugung von Nähmaschinen durch rein physikalische Prozesse weit ab
vom thermodynamischen Gleichgewicht, die Entstehung von
Kühlschränken genauso planlos im Zuge der Eiszeit etc. - und
dann, wenn es um den naturwissenschaftlichen Beweis, die
Reproduzierbarkeit der behaupteten Ereignisse geht und absolut nichts mehr
klappt, die Behauptungen mit "der Wirkung der Zeit" gegen jeden
vernünftig-realistischen Widerlegungsversuch total immunisieren.
Nun ist die Wahrscheinlichkeitsstruktur der Ereignisse
und Veränderungen, die sich in großen Zeiträumen
abspielen, sogar weitgehend bekannt: Wie oben ausgeführt, haben wir
drei Zeitraffer in der Hand, - und dennoch keine Evolution zu
primären neuen Arten (siehe auch die Wahrscheinlichkeitsansätze
J.F.Coppedge, S.Pullen [siehe LINK: Evolution durch Gen-Duplikationen?]
und von Klaus
Wittlich [siehe Literatur]). "Wenn 1000 x 0 = 0 ist, so kann,
überspitzt formuliert, nur ein Neodarwinist erwarten, dass bei der
Multiplikation von 100 000 000 x 0 mehr herauskommt" (W.-E.L.,
1. Staatsexamensarbeit, p. 86).
___
1 Von mir ergänzt.
# W.R. Bird bemerkt zum Begriff Naturgesetz 1987, pp. 24, 29,
218, 242, 243, in seiner Arbeit THE ORIGIN OF SPECIES REVISITED (Vol. II; Philosophical Library New
York) u.a. "Interestingly enough, the concept of natural
laws
originated in theology:
Bacon, for example, wrote that "the laws of Nature, which now remain and govern inviolably till the end of the
world, began in force when God first rested from his works and ceased to create; but received a revocation
in part by the curse, since which time they change not." ...In fact, the historical
origin of the natural laws concept was in creationist beliefs, according to Jaki, Whitehead, Feuer, Gilkey, and
others discussed in Section 13.4. ...As Eiseley says, "it is surely one of the curious paradoxes of the
history of science, which
professionally has little to do with faith, owes its origins to an act of faith that the universe can be
rationally interpreted, and that science today is sustained by the assumption. ...A. Whitehead,
Science and the Modern World 12-13 (1925) ("It must come from the medieval insistence on
the rationality of God, conceived as with the personal energy of Jehovah and with the
rationality of a Greek philosopher...My explanation is that the faith in the possibility of
science, generated antecedently to the development of modern scientific theory, is an
unconscious derivative of medieval theology.") .....Hodgson, Book review of Jaki's Science and
Creation, 251
Nature 747
(1974)("Although we seldom recognize it, scientific research requires certain basic beliefs about the order and
rationality of matter, and its accessibility to the human mind...They came to us in their full force through the
Judeo-Christian belief in an omnipotent God, creator and sustainer of all things.")"
##(a) Von manchen Autoren wird das Adjektiv
"heterozygot" (griech. heteros: verschieden; zygon: Joch [zygotos: zusammengejocht]) nur auf
Organismen und Zellen angewandt. Thain und Hickman definieren jedoch: "heterozygous - Designating a locus,
or organisms, at which the two representatives (alleles) in any diploid cell are different. Organisms are
sometimes described as heterozygous for a character determined by those alleles, or heterozygous at
the locus concerned. Thus the two alleles A and a occupy the A-locus, of the three genotypes
possibe (AA, Aa, aa), Aa is heterozygous while the other two are
HOMOZYGOUS" (The Penguin Dictionary of Biology. Tenth Edition 2000, p. 206; Schriftbild von den Verfassern). -
Rieger/Michaelis/Green schreiben zum Begriff Allel unter anderem: "With respect to a given locus a
diploid may be homo- (two identical alleles in the pair of chromosomes) or heterozygous (two different
alleles). If the phenotype of a heterozygous allele pair resembles that of the homozygous pair (A/A), A
is said to be dominant, a recessive" (Glossary of Genetics and Cytogenetics 1976, p. 17 [bold von
mir]. Springer, Berlin.
(b) Ich hatte zuvor von "nichtgekoppelten (mutanten) Allelen" gesprochen. H. Nilsson bemerkt
jedoch: "Ob die
Gene frei spalten oder Verkoppelung zeigen bedeutet dabei nichts. Das kann die Frequenz der Varianten verschieben,
aber auch die selteneren Rekombinanten müssen immer bei der sehr großen Individuenzahl der Arten in dem
Naturexperiment hervorgehen" (Synthetische Artbildung 1953, pp. 248/249).
*Und nach den 'allerneuesten' Berechnungen
geht
die Zahl wieder
hoch: "Haseltine, one of the most respected - and wealthiest - geneticists
of his generation, has a guess: 120,000. He said the world's best genetic
minds missed tens of thousands of genes in their haste to produce the
completed map" - Boston Globe Online, 9. 4. 2001.
**Im Sinne des morphologisch-ethologischen
Artbegriffs, genetisch hingegen sind bisher alle untersuchten Formen
miteinander fertil, gehören also nach dem genetischen Artbegriff zu
ein und derselben Art (wahrscheinlich mehr als 1000
'Arten' mit etwa 150 'Gattungen'). "Ursprünglich ordnete man sämtliche
Arten einer einzigen Gattung zu, doch vor kurzem wurden sie aufgrund einer kladistischen
Analyse morphologischer Merkmale in zahlreiche Gattungen eingeteilt, von denen
viele in allen drei Seen anzutreffen sind" (Roger Lewin [1998, p. 83]: Die molekulare Uhr der
Evolution. Spektrum Akademischer Verlag Heidelberg; bold von mir).
***Es gibt allerdings anderenorts einige
sehr
schöne Beispiele von Verlustmutationen bei der Erbse, die für
die Züchtungsforschung brauchbar waren und in
das Kulturformensortiment Eingang gefunden haben. Das bekannteste Beispiel ist wohl die
homöotische afila-Mutante (mit Ausnahme der
Nebenblätter Verlust der Blattbildung, statt
dessen
vermehrte Rankenbildung), die heute feldmäßig angebaut
wird. Eine
ausführliche Darstellung von Verlustmutationen, die für die
Pisum-Züchtung brauchbar waren, findet der daran interessierte
Leser
bei Herbert Lamprecht (1974): MONOGRAPHIE DER GATTUNG PISUM. Graz.
- Während ich diese Zeilen schreibe, wachsen hier ganz in der
Nähe
etwa viereinhalb Millionen Erbsen der Sorte Attica, bei welcher die
Fiederblätter durch Ranken ersetzt sind, auf dem rechten Feld [von
der
Stadt aus gesehen] des Gutshofs Vogelsang am
Militärring
(sechseinhalb Hektar, pro
Quadratmeter ca. 70 Pflanzen). ****"Burbank developed more than 800 new
strains and varieties of plants, including 113 varieties of plums and
prunes, 20 of which are still commercially important, especially in
California and South Africa; 10 commercial varieties of berries; and more
than 50 varietes of lilies" (The New Encycopedia Britannica, Bd. 2, 1997,
p. 640). Trotz seiner lamarckistischen Grundhaltung, die er von Darwin
übernommen hatte, praktizierte er im wesentlichen Mendelsche
Rekombinationszüchtung.
____________________________________________
Verwandte Themen/Links:
Diskussion
von Einwänden zum Antibiotikaresistenz-Thema
Evolution
durch Genduplikationen?
Gregor Mendel: Why
his discoveries were ignored for 35 (72) years
Wolf-Ekkehard Lönnig: Artbegriff, Evolution
und Schöpfung Ein paar offene Fragen der
Evolutionstheorie sowie theologische Einwände von
Evolutionstheoretikern zum Thema Intelligent Design q) Literatur
(Vorbemerkung: Die im Anhang zitierte Literatur wird hier nicht aufgeführt.)
1a
Hartl, D.L. und Jones, E.W. (1998): Genetics (S. 61). Jones & Bartlett Publishing, Sudbury, Boston. 1b Boyer, S.H., Scott, A.F., Kunkel, L.M., Smith, K.D. (1978): The proportion of all point mutations which are unacceptable: an estimate based on hemoglobin amino acid and nucleotide sequences. Canadian Journal of Genetics and Cytology 20, S. 111-137 (Zitat S. 111). 1c Schulz, L.-C.: Krankheitsmechanismen als Überlebenshilfe in den Fährnissen der Evolution. Naturwiss. Rundschau 42 (1989), S. 169-179. - 2 Muller, H. J.: Die Steuerung der menschlichen Evolution. In: L. Weß (Hrsg.): Die Träume der Genetik. Nördlingen 1989, S. 182. - 3 Beardsley, T.: Punctuated Equilibrium: Darwin survives as the debate evolves. Scientific American. März 1990, S. 16-17. - 4 Monod, J.: Zufall und Notwendigkeit. München 1971. S.211. - 5 Sitte, P.: Morphologie. In: E. Strasburger: Lehrbuch der Botanik für Hochschulen. 33.Auflage, Stuttgart 1991, S.159 und in der 34. Auflage von 1998, S.144. - 6 Ebenda 1991, S.493 und 1998, S. 479. - 7 Vgl. Lönnig, W.-E.: Artbegriff, Evolution und Schöpfung. 3. Auflage, Köln 1993, S.572-583 (1. Auflage 1986 unter dem Titel Artbegriff und Ursprung der Arten). - Kunze, R., H. Saedler and W.-E. Lönnig: Plant Transposable Elements. In: Advances in Botanical Research 27, pp. 331-470 (1997). Lönnig, W.-E., Saedler, H. : Plant transposons: contributors to evolution? Gene 205, 245-253 (1997). Lönnig, W.-E. : Natural Selection. In: The Corsini Encyclopedia of Psychology and Behavioral Sciences. Third edition. Vol. 3, S. 1008-1016. Edited by W.E. Craighead und C.B.Nemeroff. John Wiley & Sons. New York 2001. - 8 Monod, J. (Anm.4), S. 141-142. - 9 Lönnig, W.-E.: (Anm.7, Artbegriff), S. 325-463. - 10 Zur Kritik der Endosymbionten-Hypothese siehe Junker, R.. /Scherer, S. (Hrsg.): Ents tehung und Geschichte der Lebewesen. Gießen 1992. S. 122-124; und Schneider, H.: Stammen die "Kraftwerke" unseres Körpers von unverdauten Bakterien ab? In: G. Meskemper (Hrsg.): Ansätze zu einem neuen Denken. Neuhausen-Stuttgart, 1985, S. 58-107. - 11 Lönnig, W.-E.: (Anm.7, Artbegriff), S. 1-622. -11a Simmonds, N.W.:Principles of Crop Improvement. London 1979, S. 192. - 12 Leibenguth, F.: Züchtungsgenetik. Stuttgart 1982, S.207-208. Vergleiche auch Gottschalk, W.: Allgemeine Genetik. 3. Auflage, Stuttgart 1989, S. 206. - 13 Micke, A.: Utilization of induced mutants as hybrids. Genetica Agraria 22 (1970), S.262-268. - 14c und 15 Fischbeck, G./Röbbelen, G./Stutzer, D.: Landwirtschaftliche Pflanzenzüchtung in Deutschland. Herausgegeben vom Bundesverband Deutscher Pflanzenzüchter e.V. Bonn. Gelsenkirchen-Buer 1987, S. 33. -14 Auskunft eines Leiters privater deutscher Zuchtbetriebe. - 14a Poehlmann, J.M. (1987): Breeding Field Crops. Third Edition. Van Nostrand Reinhold. New York. 14b. Sengbusch, Peter von: Botanik. Blackwell, Hamburg 1989, S. 161. 15Fischbeck,G. /Röbbelen, G./Stutzer, D.: Landwirtschaftliche Pflanzenzüchtung in Deutschland. Herausgegeben vom Bundesverband Deutscher Pflanzenzüchter e.V. Bonn. Gelsenkirchen-Buer 1987, S. 33. - 15a Sengbusch, Peter von: siehe 14b. 16 Stubbe, H.: Genetik und Zytologie von Antirrhinum L. sect. Antirrhinum. Jena 1966, S.154. - 17Gottschalk, W.: Allgemeine Genetik. 3. Auflage, Stuttgart 1989, S.180. - 18 Siehe auch Blixt, S. /Gottschalk, W.: Mutation in the Leguminosae. Agri Hort. Genet. 33 (1975), S.33-85. - 19 Lundqvist, U.: Barley Mutants Mutants - Diversity and Genetics. In: G. Olsson (Hrsg.): Svalöf 1886-1986, Research and Results in Plant Breeding. Stockholm 1986, S. 76-84. - 20 Persönliche Mitteilung vom 30. Januar 1987. - 21 Hocket, E. A./Nilan, R.A.: Genetics. In: D.C. Rasmusson (Hrsg.): Barley. Madison/Wis. 1985, S. 187-230. - 22 Ebenda, S.191. - 23 Kuckuck, H./ Mudra, A.: Lehrbuch der allgemeinen Pflanzenzüchtung. Stuttgart 1950, S.162. - 24 Micke, A. /Weindl, K..: (ohne Titel). Mutation Breeding Newsletter 2l (1983), S.1. - 25 Gianola, D./Chapman, A.B. /Rutledge, J.J.: Responses to selection for body weight in descendants of X-irradiated rats. Genetics 91 (1979), S.627-645. - 26 Hartl, D.L.: A Primer in Population Genetics. 2.Auflage, Sunderland/Mass. 1988, S. 275. 26a Hartl, D.L. & Jones, E.W.: Genetics. Principles and analysis. Fourth Edition. Sudbury, Massachusetts (Jones and Bartlett Publishers) 1998, S. 686. - 26b(1)Einhorn, D.: Erfahrung und Deszendenztheorie (Verlag W. Braumüller). Wien und Leipzig 1924. - 26b(2) Zitiert nach Bird, W. R.(1989): The Origin of Species Revisited: The Theories of Evolution and Abrupt Appearance. Vol. 1. New York (Philosophical Library), 1989, S. 85.- 26c Mayr, E.: Eine neue Philosophie der Biologie. Piper, München 1991, S. 43. - 26d Mayr 1988, pp. 19-20; zitiert nach W.J. ReMine: The Biotic Message. Evolution versus Message Theory. Saint Paul 1993, p. 166. - 27 Blixt, S.: Mutation genetics in Pisum. Agri Hort. Genet. 30 (1972), S.l-293; hier S.4. - 28a Details vgl. Lönnig, W.-E.: Auge widerlegt Zufalls-Evolution. 2.Auflage, Köln 1989; und Wittlich, K..: Über die Wahrscheinlichkeit der zufälligen Entstehung brauchbarer DNA-Ketten. In: W.-E. Lönnig: Zehn Paradebeispiele gegen Zufalls-Evolution. Köln 1991, S.2 28-38. Zum Thema Genduplikationen vgl. auch: Lönnig W.-E.: (Anm.7, Artbegriff), S.440 f. - 28b Lönnig: Natural Selection; in: The Corsini Encyclopedia of Psychology and Behavioral Sciences. Third Edition. Vol 3, p. 1011. (Edited by W.E. Craighead and C.B. Nemeroff)John Wiley & Sons; New York (2001) - 28c Remane, A.: Offene Probleme der Evolution. In: Evolution (Nova Acta Leopoldina; Hrsg. J.-H. Scharf, Nummer 218, Band 42). Halle, Saale 1975, S. 169. - 28d Kühn, A. Grundriß der allgemeinen Zoologie. Stuttgart 1967, S. 371/372. - 28eDenton, M.: Nature's Destiny. The Free Press. New York 1998. - 28fKlekowski, E.J.: Mutation, Developmental Selection, and Plant Evolution. Columbia University Press. New York 1988. - 29 Vavilov, N.I.: The law of homologous series in variation. J. Genet. 12 (1922), S.47-89. - 30 Haecker, V.: Pluripotenzerscheinungen. Jena 1925, S.7. - 31 Ebenda, S.9. - 32 Lamprecht, H.: Monographie der Gattung Pisum. Graz 1974. 32aIch hatte zunächst angenommen, dass bisher überhaupt keine "geflügelte" Mutante von der Erbse beschrieben worden sei. Inzwischen habe ich beim Durcharbeiten älterer Literatur scheinbar(?) eine Ausnahme gefunden. Eriksson erwähnt 1929 eine Mutante, die er nach Kreuzungen verschiedener Erbsensorten erhalten hat (p. 446): "...gewisse Linien in der Bastardierung Stens x Concordia [haben] gewisse mißbildete Individuen gegeben. Diese Deformationen waren folgende: 1. In F2 eine Pflanze mit geflügeltem Stengel und langschmalen Blättern. Lathyrustypus mit allen Blumenteilen in langschmale Blätter umgewandelt." Leider hat Eriksson keine genauere Beschreibung mit einer Abbildung - das wäre das Wichtigste - präsentiert. Ich hätte zu gern gewusst, ob tatsächlich typische Lathyrusmerkmale in der Stengelausbildung vorhanden waren, oder ob vielleicht mehr eine Art Fasziation vorgelegen hat. Jedenfalls war die Pflanze total steril: "alle Blumenteile in langschmale Blätter umgewandelt" (also keine Staubblätter und keine Fruchtblätter vorhanden, d.h. keinerlei Fortpflanzungsorgane, weder männlich noch weiblich, statt dessen nichts weiter als laubblattähnliche Blattbildungen) (Eriksson: Z. Pflanzenzüchtung 14:445-476, 1929). 33 Ayala, F.J./Kiger, J.A.: Modern Genetics. 2.Auflage, Menlo Park/Cal. 1984, S.678. - 34 McKusick, A.V.: Mendelian Inheritance in Man. 8.Auflage, Baltimore 1990. - 35 Schmidt, F.: Grundlagen der kybernetischen Evolution. Krefeld 1985, S.26, 31. - 36 Nilsson, H.: Synthetische Artbildung. Lund 1953, S.251-253. - 37 Vogel, F./Motulsky, A.G.: Human Genetics. 2.Auflage, Berlin 1986, S.636, 641. - 38 Gottschalk, W. (Anm. 17), S. 313. - 39 Strickberger, M. W.: Genetik. 3. Auflage, München/ Wien 1988 (deutschsprachige Ausgabe), S.117. - 40 Zimmermann, W.: Methoden der Phylogenetik. In: G. Heberer (Hrsg.): Die Evolution der Organismen. 2.Auflage, Stuttgart 1959, S.25-102. - 41 K. (Ich gebrauche hier diese Abkürzung, da ich den Eindruck hatte, das Herr Prof K. zunehmend unglücklich über unsere Diskussion war. Mir geht es jedoch um die Sache, nicht darum jemanden persönlich anzugreifen oder herabzusetzen (ich gebe K.'s Einwände hier wieder, weil ich sie für repräsentativ für den Neodarwinismus halte). Natürlich ist es nicht zu ändern, dass man die Identität auf Grund der schon publizierten Arbeiten leicht herausfinden kann): Neodarwinsche Theorie und Makroevolution: Probleme antidarwinistischer Kritik. Biologie heute 381 (12/1990), S.8. (Zu den weiteren Einwänden siehe auch: Lönnig, W.-E.: Kann der Neodarwinismus durch biologische Tatsachen widerlegt werden? Köln 1991.) - 42 Ebenda, S. 8. - 43 Ebenda, S.8. - 44 Ebenda, S.8. - 45a Ebenda, S.8. - 45b Margulis, L., Sagan, D.: Slanted Truths: Essays on Gaia, Symbiosis, and Evolution. Springer-Verlag, Berlin 1997, S. 265 und 279 (die Zitate zwischen den beiden längeren Passagen stammen von S. 266, 270, 272, 275) 45c Mann, C.: Lynn Margulis: Science's unruly earth mother. Science 252, S. 378-381 (1991).46a Ernst Mayr (1942): Systematics and the Origin of Species. Columbia University Press, New York, S. 296. 46b Strugger, S.: Botanik. Stuttgart 1962, S.59. 46c Die Literaturhinweise implizieren nur meine Identifikation mit den zutreffenden naturwissenschaftlichen Argumenten, in der Regel jedoch nicht mit den (eventuell) weiteren weltanschaulichen Anliegen der Verfasser. - 47 Ausführliche Begründungen in Lönnig, W.-E.: (Anm.7, Artbegriff), S.34,50,56. - 48 Hier gäbe es allerdings als weitere Alternative die kontinuierliche Schöpfung, die diskontinuierliche aber wäre widerlegt.