
1. 1. WIRKUNG DER GENMUTATIONEN
Der Genetiker und Nobelpreisträger Muller meinte 1946, p. 38: "Most mutations are bad. In fact, good ones are so rare that we can consider them all as bad." Abgesehen von den etwa 20 % Basenaustauschmutationen, die wegen des degenerierten Codes ein neues Gleichsinncodon bilden, führen die meisten der übrigen 80 % zu Funktionsausfällen - ein Punkt, der in der gesamten älteren und neueren Literatur immer wieder betont wird und den ich in der vorliegenden Arbeit schon auf den Seiten 122-137 erwähnt und mit den Rekombinationsquadraten veranschaulicht habe. Fehlsinncodons sowie vorzeitiger Abbruch der Translation eines Polypeptids und Rastermutationen etc. werden in der Regel nicht zur Verbesserung des Codes führen. In der Praxis sieht die Situation so aus, wie sie von Esser und Kuenen in ihrem Buch über die Genetik der Pilze (1965) beschrieben wurde. Die Liste dürfte heute nur wesentlich länger sein und detaillierte Angaben über qualitative Änderungen enthalten. Die Autoren schreiben unter anderem (p. 381):
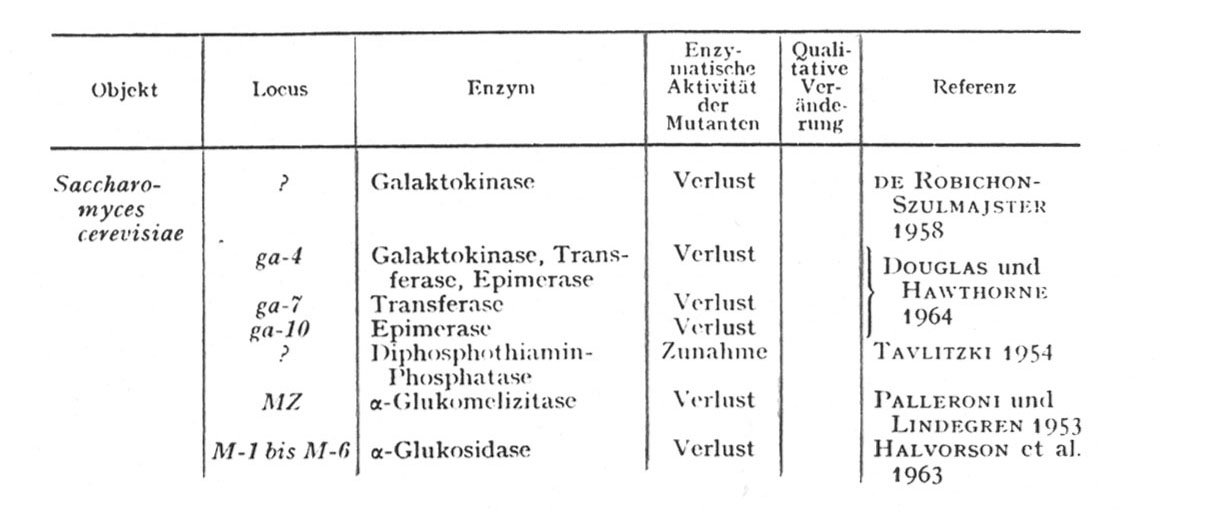
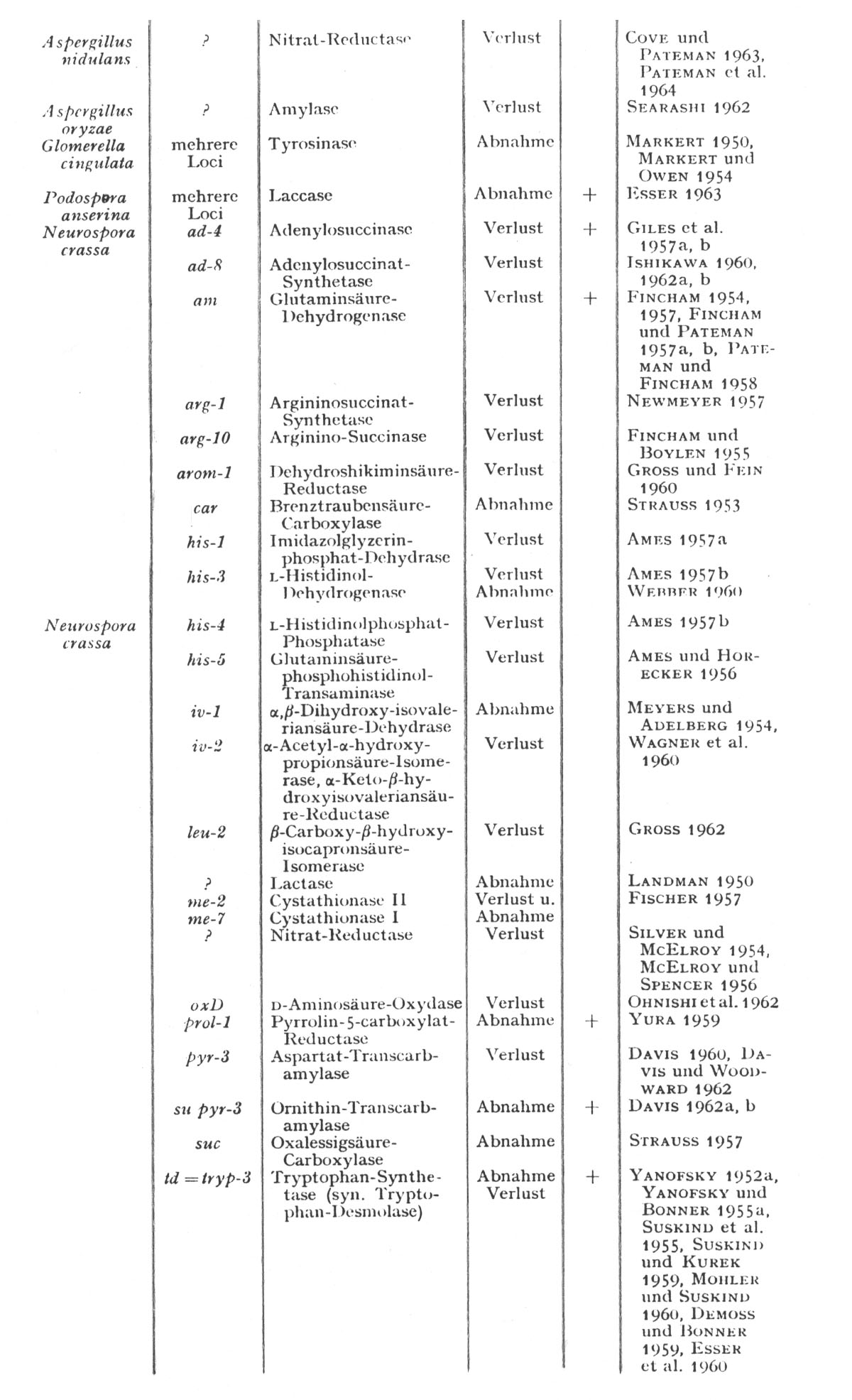
Der am häufigsten beobachtete quantitative Effekt einer Genmutation führt zu einem Ausbleiben der Enzymbildung bei der betreffenden Mutante. In anderen Fällen besteht die Auswirkung der Mutation in einer Veränderung der gebildeten Enzymmenge. Meistens war die relative Aktivität herabgesetzt, und nur in einem Fall (TAVLITZKI 1954) ergab sich bisher eine Zunahme (Tabelle VI-3).
Die von Esser und Kuenen zusammengestellte Tabelle zu diesem Thema sei im folgenden wiedergegeben (pp. 382/383):
Tabelle 6 (Aus Esser und Kuenen 1965): "Beispiele für enzymatische Veränderungen in Mutanten verschiedener Pilzen: Mit Ausnahme der prototrophen Tyrosinase- und Laccase-Mutanten sind alle übrigen Mutanten auxotroph. Mutanten-Enzyme, für die mit Sicherheit strukturelle Veränderungen nachgewiesen wurden, sind mit + gekennzeichnet. Alle anderen Enzyme sind entweder nur quantitativ verändert oder noch nicht hinreichend auf qualitative Änderungen untersucht worden (unter Verwendung der Angaben von FINCHAM und CATCHESIDE 1960a)."
Mir ist kein Fall bekannt, in dem Genmutationen zum Aufbau eines völlig neuen Enzyms geführt hätten, so dass die Enzymlisten aufgrund von Punktmutationen bei den verschiedenen Arten erweitert werden müssten. (Siehe auch Nachtrag p. 545.) Auf die seltenen Fälle von Zunahme der Enzymaktivität kommen wir p. 365 zurück. Die von O'Brien (1980, 1982, 1984) herausgegebenen Genkarten führen zwar nur die chromosomal lokalisierten Gene auf, die dazu zitierte Literatur gibt jedoch in der Regel einen umfassenden Überblick über das gesamte Mutationsspektrum der betreffenden Arten (mehr als 50 Arten in Bd. 3). Somes hat 1980 die Genliste des Haushuhns zusammengestellt, aus der ich im folgenden die ersten Buchstaben zitiere:
Tab. 7 (aus Somes 1980): "Alphabetical listing of chicken loci and alleles"
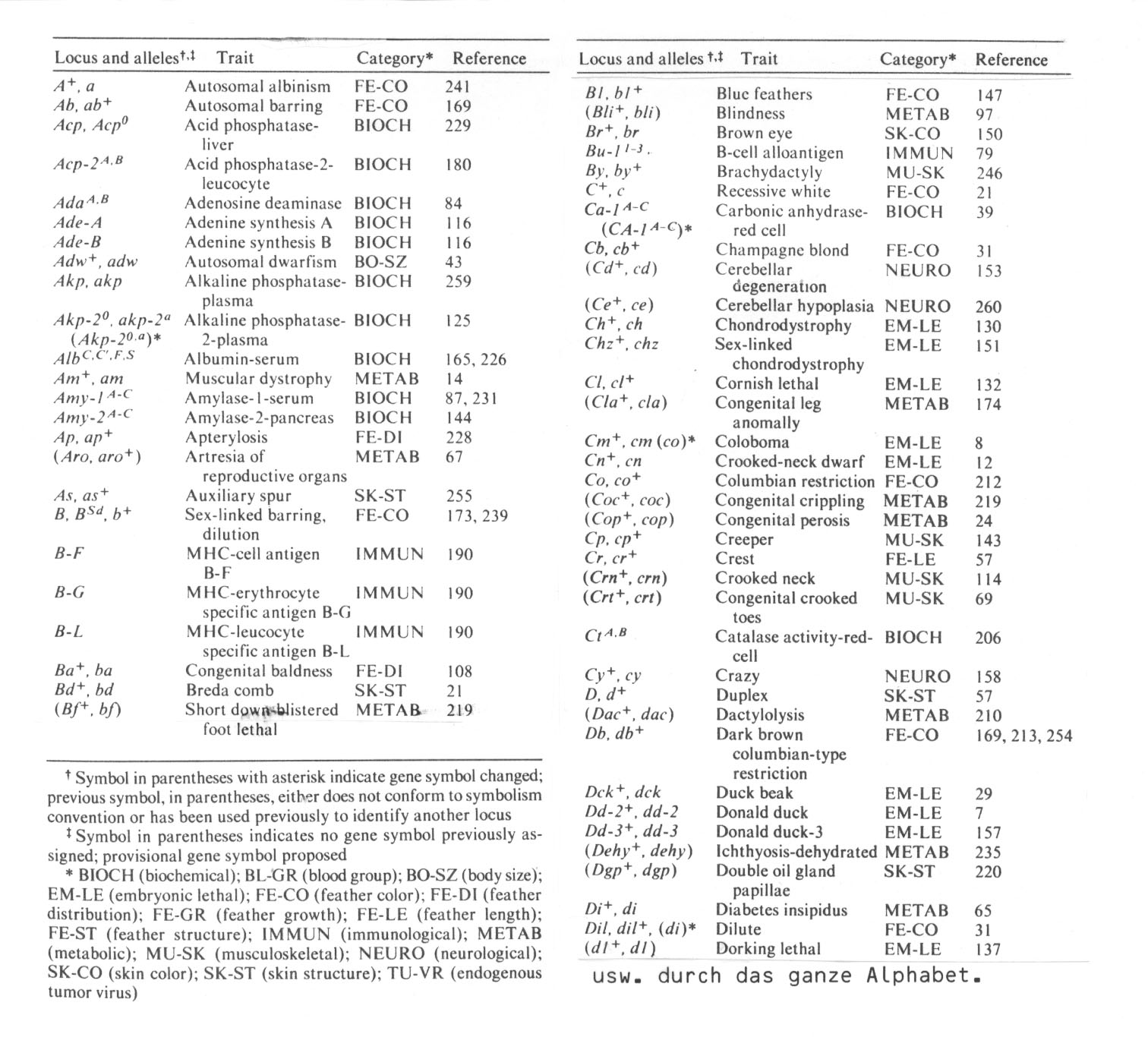
Bei den unter BIOCH (biochemical) aufgeführten Genloci handelt es sich desöfteren nicht um durch spontane Mutationen erhaltene Daten, sondern um (gelelektrophoretische) Nachweise von Enzymen der Adeninsynthese, von Plasmaalbuminen, Isozymen der Pankreasamylase-2 etc. (z.T. nach Zellfusionsexperimenten).
Die Mutationen in Genen für Farbstoffsynthesen führen (zumindest in der Haustierzucht) häufig zu neutralen Allelen. Es leuchtet ein, dass die meisten Mutationen, die nur die Farbe des Federkleides betreffen, nicht unmittelbar lebensgefährlich zu sein brauchen. Dennoch handelt es sich auch hier in der Regel um Strukturabbau: Autosomal albinism, autosomal barring, recessive white etc. (vgl. die ähnliche Situation bei Pflanzen und das Prinzip der Variabilitätserhöhung durch Funktionsausfall pp. 193/194).
Für die betroffenen Individuen wesentlich schwieriger wird die Situation schon bei Mutationen in der Federstruktur und -verteilung sowie im Federwachstum: Bei der Apterylosis z.B. handelt es sich um 'nakedness due to absence of pterylae' mit stark schwankender Expressivität. Zum Selektionswert von Muskeldystrophie, 'Short down blistered foot lethal', Blindheit, Brachydaktylie, Cerebellum-Degeneration, Angeborener Verkrüpplung etc. und allen EM-LE (embryonic lethal) Mutanten braucht man nichts hinzuzufügen. Interessanterweise tauchen gerade bei der letzteren Gruppe ein paar morphologisch interessante Mutanten auf wie "Duck beak" u.a.. Es wird sich jedoch mit Sicherheit auf diesem Wege keine Ente vom Huhn ableiten lassen, zumal die Entenfamilie nicht nur durch den besonderen Schnabel (der selbstverständlich bei den EM-LE Mutanten nur an den Entenschnabel erinnert, nicht aber entenspezifisch ausgebildet wird), sondern durch die Synorganisation einer ganzen Kette weiterer morphologischer Merkmale und Verhaltensweisen (Hornlamellen im Schnabel, Ruderfüße mit Schwimmhäuten, Begattungsglied beim Männchen, dichtes Gefieder, gutes Flugvermögen, ethologische Programme wie Neigung, schon als Küken zum Wasser zu gehen und zu schwimmen etc.) auszeichnet.
Kein Biologe und Züchter erwartet aufgrund dieser reichlich vorhandenen Mutationsdaten und allen daraus folgenden Rekombinationsmöglichkeiten jemals aus dem Haushuhn eine systematisch völlig neue Tiergruppe züchten zu können.
Werfen wir zu diesem Thema einen Blick auf das mutationsgenetisch am gründlichsten untersuchte 'Haustier': Aus der Drosophila-Forschung lagen nach der Literaturübersicht von Ashburner und Novitzki schon 1976 etwa 25 000 Einzelarbeiten vor, zum größten Teil mit dem Schwerpunkt Mutationsgenetik. Von einer durch mutagene Agenzien und/oder durch Rekombination verschiedener mutierter Gene hervorgerufenen neuen Drosophila-Art ist jedoch nirgends die Rede.
Wenden wir uns der Humangenetik zu:
McKusick bringt die Genliste des Menschen laufend auf den neuesten Stand und wir geben im folgenden seine Tabelle aus GENETIC MAPS (1984 p. 455) wieder:
Tab. 8 (aus McKusick 1984): "Number of loci identified mainly by mendelizing phenotypes*. From McKusick: Medelian Inheritance in Man, 6th ed., 1983."
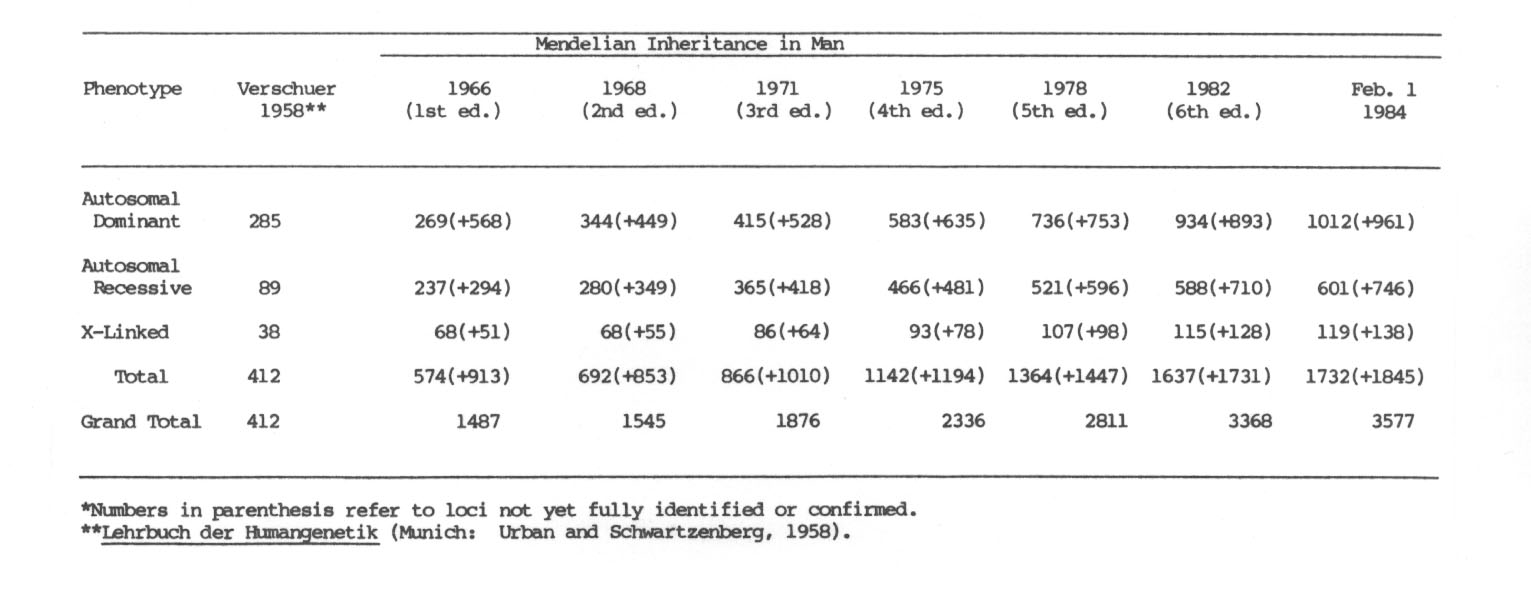
Die meisten dieser Gene sind aufgrund von Genmutationen nachgewiesen worden.
Schmidt schreibt zu den von McKusick aufgeführten erblichen Abweichungen (1985, p. 26): "Fast alle davon sind als Krankheiten oder Mißbildungen im medizinischen Sinne des Wortes anzusprechen." - Und er fährt auf der Seite 31 fort:
Gewiß fallen Mutationen, die schwere körperliche und geistige Schäden verursachen, mehr auf und stellen demnach eine gewisse Interessantheitsauslese dar, weil die betreffenden Patienten früher oder später den Weg zum Arzt finden. Die Tatsache aber, daß Hunderte von Genetikern, die im letzten halben Jahrhundert genetische Untersuchungen am Menschen durchführten, in keinem einzigen Falle der Nachweis einer überzeugenden vorteilhaften Mutation gelang, ist ein klarer Beweis dafür, wie unsicher das Fundament ist, auf dem der Neodarwinismus einen wahren Wolkenkratzer an Spekulationen errichtet hat.
Zur Veranschaulichung der von McKusick aufgeführten Mutationen sei die Seite 1275 [Das ist die 1. Seite der zusammenfassenden Liste aller bis dahin bekannten mendelnden Eigenschaften beim Menschen.] der Liste aus seinem Werk MENDELIAN INHERITANCE IN MAN (1983) wiedergegeben:
Tab. 9 (aus McKusick 1983):


Kein Forscher erwartet aufgrund der beim Menschen bekannt gewordenen Mutationen etwa die "Entwicklung neuer Menschenarten". Man befürchtet vielmehr erhöhte Mutationsraten durch die zunehmende Umweltbelastung (chemische und strahlenbedingte Mutagenese) und arbeitet z.B. an der Herabsetzung von MAK-Werten beim Umgang mit mutagenen Agenzien, zumal die meisten dieser Agenzien auch noch karzinogen sind. An dieser Stelle darf man wohl die Frage nach der Berechtigung der Behauptung stellen, dass der Mensch durch die uns bekannten Mutationen ins Dasein gekommen ist. Wie ist das möglich, wenn diese Mutationen für eine Weiterentwicklung untauglich sind?
Zur Mutationsfrequenz beim Menschen schreiben Ayala und Kiger 1980, pp. 550/551 (vgl. auch 1984, p. 678; diese Werte sind natürlich für die jeweils neuesten Zahlen zur Weltbevölkerung hochzurechnen):
Assume that there are 100 000 pairs of genes in humans...and that the average mutation rate per gene per generation is 10-5. The average number of mutations arising per generation can then be estimated as (2 x 105 genes) x (10-5 mutation per gene) = 2 mutations for a human zygote...There are about 4 x 109 humans in the world. With two mutations per individual, the total number of mutations newly arisen in the present population becomes 8 x 109.
........
When whole species are considered, many mutations occur in every generation, even at a single locus. If the average mutation rate per gene per generation is 10-5, about 80 000 new mutations arise, on the average, per generation at each locus in the human species (4 x 109 individuals x 2 genes per locus x 10-5 mutation per gene).
Eine Aufführung ähnlicher Berechnungen verschiedener Autoren findet sich bei Schmidt (1985, pp. 30 - 54). [S. Borojevic rechnet mit 1000 Mutationen/Locus*/Tag/Hektar für Puccinia recondita (leaf rust in wheat). Man errechne einmal die Zahl der Mutationen weltweit! (Principles and Methods in Plant Breeding; Elsevier 1990, p. 32)]
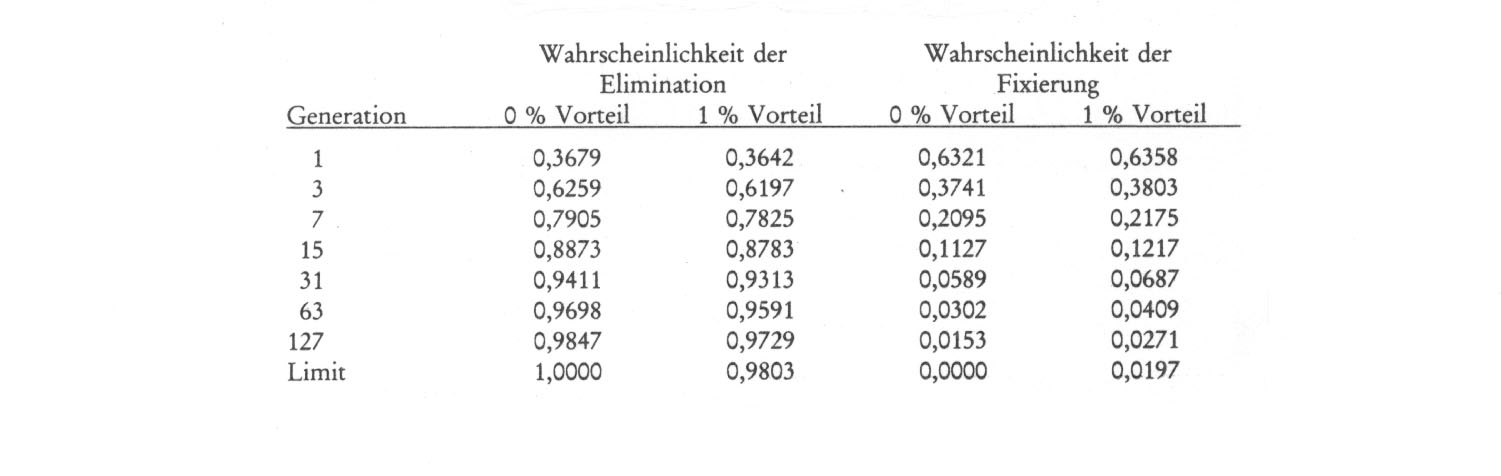
Wenn nun jedes einzelne Gen pro Generation in der zitierten Population etwa 80 000 Mal mutiert, dann darf man wohl ruhigen Gewissens feststellen, dass in solchen Fällen in jeder Generation fast das gesamte Punktmutationspotential der Art durchgespielt und (zum Teil auch in Verbindung mit der Rekombination) freigesetzt wird. Beim Menschen wirken sich im Gegensatz zu den bekannten Daten aus der Pflanzengenetik die mutierten Allele von fast zwei Dritteln der festgestellten Loci dominant aus (vgl. die Angaben von McKusick, zitiert p. 338) und sind damit schon beim Auftreten phänotypisch festzustellen. Dennoch entwickeln sich nirgends neue Spezies aus dem Homo sapiens, und wie wir schon feststellten, ist das bei der bekannten 'Qualität' aller Mutationen auch nicht zu erwarten. Aber selbst Mutationen mit einem Prozent Selektionsvorteil würden im Laufe der Zeit meist wieder verloren gehen. Schmidt zitiert (1985) dazu die Tabelle zur Wahrscheinlichkeit der Eliminierung oder Fixierung einer Mutation in einem Individuum mit einem postulierten Selektionsvorteil von 0 % und 1 % nach Fisher:
Tabelle 10 (nach Fisher und Dobzhansky aus Schmidt 1985): "Wahrscheinlichkeiten für die Eliminierung oder Fixierung einer Mutation in einem einzelnen Individuum bei einem angenommenen Selektionsvorteil von 0 % und 1 %."
- Und fügt hinzu:
Aus der Tabelle geht hervor, daß die Wahrscheinlichkeit der Eliminierung einer Mutation sowohl ohne Selektionsvorteil, als auch bei 1 % Vorteil schon nach 31 Generationen weit über 90 % erreicht.
(Vgl. auch Kimura, zitiert p. 240)
Weiter bemerkt Schmidt (1985, p. 47) zu solchen eventuell vorteilhaften Mutationen:
Selbst wenn eine solche Mutation durch einen ganz außergewöhnlichen Zufall vielleicht einzelne Generationen überdauert, geht der etwaige Selektionsvorteil unter der sehr viel größeren Zahl neu auftretender Defektmutationen unter und könnte somit der Selektion nur in Ausnahmefällen einen Ansatzpunkt bieten. Die natürliche Zuchtwahl wäre im Gegenteil voll damit ausgelastet, diese neu hinzukommenden nachteiligen Mutationen zu eliminieren. In jedem Falle muß die Gesamtbilanz - auch wenn man grundsätzlich die Möglichkeit von Gewinnmutationen anerkennt - durch die gleichzeitig auftretenden, sehr viel häufigeren Defektmutationen speziell bei höheren Arten negativ sein. Das beweist: Auch die natürliche Zuchtwahl kann den Neodarwinismus nicht aus diesem Dilemma herausführen.
Unter diesen Umständen gehört ein wahrhaft übermenschliches Maß an Optimismus dazu, der neodarwinistischen Interpretation der Evolution zuzustimmen. Man könnte die Situation mit einer Fabrik vergleichen, in der auf jeden Beschäftigten, der sich bemüht, durch eine Neuerung (= Gewinnmutation) die Produktivität zu steigern, mindestens 100 Saboteure kommen, die jeden unbeobachteten Augenblick benutzen, um Sand in das Getriebe zu streuen (= Defektmutation). Unter diesen Umständen einen florierenden Produktionsbetrieb (= Evolution) zu erwarten, ist wohl kaum realistisch.
Demnach ist die Mutationsfrequenz so hoch, dass die Gesamtbilanz selbst dann noch negativ ist, wenn man 1 % Mutationen mit positivem Selektionswert ansetzt (dieser Selektionswert ist nach allen bekannten Daten sicher zu hoch angesetzt; selbst ein Optimist zur Entstehung der Arten durch Mutation und Rekombination wie J. Huxley rechnete nur mit 1 Promille positiver Mutationen, und auch dieser Wert ist mit Sicherheit noch zu hoch - vgl. p. 351 f.).
Das erklärt uns auch die allgemeine Tendenz zur Degeneration in der Organismenwelt. Außerdem kann Selektionsvorteil auch mit Strukturabbau (flügellose Insekten der Kerguelen und anderen ozeanischen Inseln) bzw. Rückmutationen verbunden sein. Mit den obigen Rekombinationsquadraten (pp. 122 f.) und der Diskussion zum genetischen Artbegriff haben wir schon im Detail veranschaulicht, dass unter verschiedenen ökologischen Bedingungen Mendelsche Rekombination zwar mit unterschiedlichen Selektionswerten verbunden sein kann, dass diese Frage jedoch nichts mit primärer Artbildung zu tun hat.
Interessanterweise wird auch von der Synthetischen Evolutionstheorie die Hybridisation als Ursache der Artbildung für das Tierreich einhellig abgelehnt und für das Pflanzenreich auf die Bildung von Morphospezies (= Populationen ähnlicher Rekombinanten) und die Polyploidie beschränkt (Literatur bei Grant 1981, pp. 202 - 204).
Da nach dieser Theorie alle Gene und Allele durch Mutation entstanden sind und "die Evolution im allgemeinen Sinne der Genetik nichts anderes als Genfrequenzänderungen beinhaltet" (Köhler und Belitz 1976, p. 263) (und somit sich durch Bildung und Rekombination neuer Allele Unterarten zu Arten, Arten zu Gattungen, Gattungen zu Familien etc. weiterentwickeln sollen), müsste die Hybridisation die ideale Methode zur experimentellen Erzeugung auch höherer systematischer Kategorien sein. Bei Artkreuzugnen mit fertiler Nachkommenschaft müssten bei einer großen Population schon in der F2 neue Arten und Gattungen herausspalten und die Nachkommenschaft fertiler Hybriden aus Gattungskreuzungen sollte sogar zu neuen Familien führen. Denn nach den Voraussetzungen der Theorie werden durch Hybridisation große Zahlen mutierter (und im selektionistischen Sinne erprobter) Gene in der F1 zusammengeführt und durch die Rekombination dauerhaft vereinigt. Mit 100 hinzukommenden heterozygoten Allelpaaren** wird mit einem Schlag das Rekombinationspotential für mindestens 1 267 650 600 000 000 000 000 000 000 000 phänotypisch nachweisbaren Rekombinanten geschaffen. D.h. der weite Weg eines neu mutierten Gens aus dem heterozygoten Stadium durch schrittweise Anreicherung in der Population, wiederholter mutativer Umstrukturierung zur Fitnesserhöhung, Homozygotie- und Interaktionstests in den unterschiedlichsten Genomen der Population und Erprobung unter verschiedenen ökologischen Bedingungen entfällt durch die Hybridisation, so dass auch die Bildung höherer systematischer Kategorien durch die Zeitraffermethode der Rekombination direkt beobachtet werden müsste. Wie allgemein bekannt, ist das jedoch nicht der Fall.
Der neben den neutralen und schwach nachteiligen DNA-Veränderungen oben aufgeführte Defektcharakter der meisten Punktmutationen und die damit verbundene genetische Bürde sind hingegen in voller Übereinstimmung mit den Hybridisationsergebnissen:
Durch Vereinigung funktionsfähiger Allele kann es in der F1 zu Hybrid Vigour (Heterosis) kommen und durch Rekombination funktionsloser Allele zu Hybrid Breakdown etc. (vgl. p. 124 f. ). Mit dem Ausgangspunkt maximaler Informationsdichte bei den ursprünglichen Arten und darauf folgender degenerativer Aufsplitterung in Subspezies, Rassen und begrenzte Populationen ist die Bildung neuer Arten, Gattungen und Familien durch Hybridisation auch nicht zu erwarten, zumal die Differenzen in den Strukturgenen auch nach den Recherchen einer zunehmenden Zahl zeitgenössischer Biologen (vgl. p. 212) nichts mit Artbildung zu tun haben. Die primären Arten sind hingegen durch die meiotisch-plasmatische Barriere absolut voneinander getrennt. Mit der tausendfachen empirischen Feststellung, dass durch Hybridisation keine neuen Arten, Gattungen und Familien entstehen, ist auch die Evolution durch Punktmutationen widerlegt. Artbildung ist weder durch Mutation noch durch Rekombination möglich.
Diese Schlussfolgerung wird durch zahlreiche weitere Tatsachen belegt. Die Mutationsforschung z. B. hat uns neben der ionisierenden und ultravioletten Strahlung eine Unmenge mutagener Substanzen in den letzten Jahren aufgelistet. So sind nach Maron und Ames (1983) im Environmental Mutagen Information Center Index mehr als 5 000 chemische Verbindungen aufgeführt, von denen ein großer Teil mehr oder weniger stark mutagen ist. Und diese Daten beziehen sich allein auf den Amestest (induzierte Reversionen bei Salmonella). Trotz der riesigen Zahlen induzierter Mutationen und Reversionen und der kaum mehr zu schätzenden Milliarden und Billionen von Testindividuen ist nirgends die Weiterentwicklung von Salmonella typhimurium zu neuen Arten und Gattungen etc. festgestellt worden. Dasselbe kann man für alle anderen Testmethoden mit Bakterien, vor allem für die umfangreichen und intensiven Studien an Escherichia coli feststellen. Aber auch höher organisierte Organismen wie Neurospora und die schon zitierte Drosophila sind in umfangreichen Versuchsserien für Mutagenitätstests verwendet worden. Für die letztere beläuft sich die Liste der untersuchten Substanzen auf immerhin schon mehr als 400 (genau 421, 198 sind davon positiv, d.h. mutagen, 46 negativ und bei den restlichen 177 sind die Ergebnisse statistisch nicht ausreichend genug abgesichert; - vgl. Details bei Lee et al. 1983).
_______
*Nachtrag zur Terminologie: "locus (Morgan, Sturtevant, Muller & Bridges 1915) - the position of a gene on the genetic map. Allelic genes are situated at identical loci in homologous linkage structures (chromosomes). The locus of a particular gene is identifiable as a site of an allelic difference arising by a gene mutation. Frequently the term locus is used more or less synonymously with gene..."(Rieger/Michaelis/Green: Glossary of Genetics and Cytogenetics; bold von mir). Diesem Brauch bin ich mit weiteren Autoren auch bei den obigen Ausführungen gefolgt.
**Vgl. die zweite Fußnote zum Gesetz der rekurrenten Variation.