
1. PUNKTMUTATIONEN
Vogel und Angermann kommentieren (1984, p. 473) den von Bridges 1923 stammenden Begriff der Punktmutation wie folgt: "Punktuelle DNA-Veränderung einzelner oder weniger Nukleotid(paar)e, entweder durch Basensubstitution(-ersatz) oder Rasterverschiebung läßt ein neues Allel entstehen (S. 474 f.)." Die durch Punktmutationen veränderten Basensequenzen können sich sehr unterschiedlich auf die Aminosäurensequenz des kodierten Polypeptids auswirken und die Autoren führen vier Möglichkeiten auf (p. 475):
- Ca. 20 % der Basenaustauschmutationen bilden wegen des degenerierten Codes ein neues Gleichsinnkodon und damit den Wildtyp.
- Das neue Triplett codiert über ein Fehlsinncodon eine andere Aminosäure; die Funktion des Polypeptids der Mutante kann in der Effektivität zw. 0 bis < 100 % liegen.
- Entsteht ein nichtcodierendes Stop-Codon, so bricht die Translation des Polypeptids hier vorzeitig ab; die Mutante verfügt dann meist über ein inaktives Enzym.
- Rastermutationen zerstören, wenn nicht gerade drei Nukleotide betroffen sind, die sinnvolle Information des Allels
Gottschalk unterscheidet (1984, p. 242 f.) folgende Möglichkeiten (ähnlich Dobzhansky et al. 1977, Kaudewitz 1983, Günther 1984, Knippers 1985 u.v.a.):
- Transitionen. Ersatz einer Purinbase durch die zweite Purinbase oder einer Pyrimidinbase durch die zweite Pyrimidinbase.
- Transversionen. Ersatz einer Purinbase durch eine Pyrimidinbase oder umgekehrt.
- Insertionen. Addition von Nukleotiden.
- Deletionen. Eliminierung von Nukleotiden.
Abb. 50: (aus Gottschalk 1984) Möglichkeiten mutativer Änderungen an der DNA-Doppelhelix. In jedem Fall ist jeweils links der Ausgangszustand, rechts der mutierte Zustand angegeben.
(Die Begriffe Transition und Transversion stammen von Freese 1959.)
Günther unterscheidet (1984, pp. 187 - 194) bei den Genmutationen Mikro-und Makroläsionen. Im Gegensatz zu den anderen mir bekannten Autoren, die alle Veränderungen innerhalb eines Gens als Punktmutationen klassifizieren, setzt die Autorin nur die Mikroläsionen mit den Punktmutationen gleich, die sie unter den beiden Unterthemen 'Basensubstitutionen' und 'Frameshiftmutationen' abhandelt und für welche sie generell auf ihre Abbildung 9.4. hinweist (die wir auf der nächsten Seite wiedergegeben haben). Über die beiden anderen Autoren hinaus weist sie in diesem Zusammenhang noch auf Dimerbildungen, Einzel- und Doppelstrangbrüche sowie auf Cross-links hin. Zu den Makroläsionen führt sie folgende Beispiele auf (p. 193):
Größere Veränderungen innerhalb eines Gens sind die
Duplikation: Verdoppelung einer längeren Nukleotidsequenz,
Deletion: Verlust einer längeren Nucleotidsequenz,
Inversion: Umkehr einer längeren Nucleotidsequenz um 180°,
Insertion: Einbau genfremder Sequenzen ins Gen.
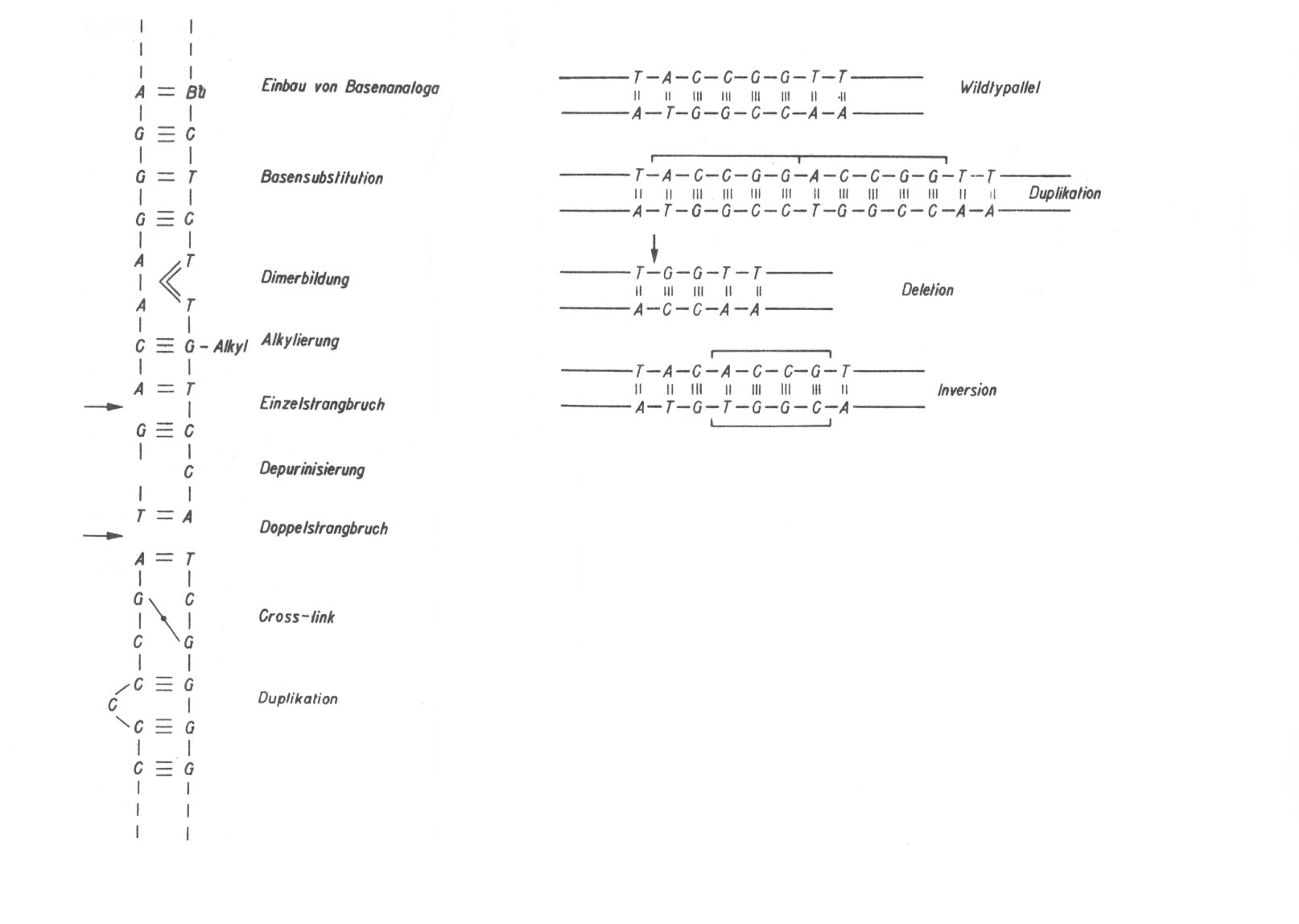
Abb. 51: (aus Günther 1984; links nach Böhme und Adler): Hier werden nicht nur die Veränderungen an der DNA gezeigt, die spontan oder nach Einwirkung mutagener Agenzien auftreten können, sondern zum Teil auch ihre Ursachen. (Rechts:) Beispiele für Makroläsionen.
Als weiteren Punkt für Veränderungen innerhalb eines Gens könnte man noch "intra-allelische" Rekombination zitieren. Kühn und Hess schreiben dazu (1984, p. 103):
Bei Mikroorganismen, vor allem Bakterien, wurde es...möglich, Nachkommen heterozygoter Individuen in sehr großer Zahl zu analysieren. Dabei stellte sich heraus, daß auch noch zwischen verschiedenen Allelen Rekombination vorkommt. Selbst bei Drosophila hat man schließlich nach Jahrzehnten intensiver genetischer Forschung gefunden, daß auch hier innerhalb eines Bereichs, den man bisher als "ein Gen" ansah, noch Austausch stattfinden kann. In seltenen Fällen (weniger als 0,1%, teilweise sogar seltener als 0,001 %) findet man deshalb in der Nachkommenschaft von Heterozygoten für zwei Allele einer Serie multipler Allele sowohl Doppelmutanten (die meist schwer zu erkennen sind) als auch (die sehr auffälligen) Wildtypen. Tatsächlich konnte man in allen bisher untersuchten Fällen nachweisen, daß Rekombination innerhalb von Genen vorkommt (die sog. "Intra-allelische Rekombination"). Durch Außenmarkierungen ließ sich zweifelsfrei zeigen, daß die Wildtypen stets nach Crossing-over auftraten, womit belegt ist, daß es sich nicht um sog. "Rückmutationen" handelt. In den homozygoten Rassen jedes der beiden Allele treten Wildtypen tatsächlich auch nicht auf. (Kursiv von den Verfassern)
Zwar werden solche DNA Veränderungen in keiner mir bekannten Arbeit unter der Rubrik der Genmutationen aufgeführt, es handelt sich jedoch auch bei diesen Beispielen (sowie bei einigen 'unequal crossingovers') um Veränderungen innerhalb des Gens, so dass ich diese Punkte der Vollständigkeit halber hier noch aufgenommen habe.
Ich habe damit einen summarischen Überblick über alle in der einschlägigen Literatur gegebenen Veränderungsmöglichkeiten innerhalb eines Gens gegeben.
Zu den Ursachen der Genmutationen führt Kornberg (1980, p. 608) folgende Punkte auf (unter Hinweisen auf die ins Detail gehenden Kapitel 11 und 12 seines Werkes) (dem an diesen Details interessierten Leser sei diese Arbeit empfohlen):
(i) Missing base. Acid and heat remove purines; at physiological temperatures 10,000 purines are lost per mammalian genome per day. Certain alterations destabilize bases, leading to their removal (see ii). Specific enzymes remove bases (e.g., uracil by an N-glycosylase).
(ii) Altered base (or nucleoside). Ionizing radiation and nucleophilic agents, including alkylating agents modify bases. Base (or nucleoside) analogs may be substituted during replication.
(iii) Incorrect base. Improper base pairing may result from an incorporation error in replication or from deficient proofreading. Mutator genes may code for defective polymerases, nucleases, or other proteins essential for high-fidelity replication. Deaminations (cytosine to uracil, adenine to hypoxanthine) are both spontaneous, because of inherent chemical instability, or inducible by nitrous acid. Mutations due to deamination are predictably not transversions but transitions. That is, one pyrimidine replaces the other, or one purine replaces the other; for example, TA becomes CG. (In transversions a purine replaces a pyrimidine, a pyrimidine replaces a purine; e.g. TA->AT.)
(iv) Deletion or insertion. Intercalating agents (e.g. acridines), inserted between the planar rings of the bases, induce replication or recombination errors by omission or addition of an extra base. These errors produce frameshift (phase-shift) mutations, which put the reading frame for translation of mRNA out of phase from the point that the nucleotide (or two) was inserted or deleted.
(v) Cyclobutyl dimer. Ultraviolet irradiation (254 nm) produces dimer adducts between adjacent pyrimidine bases, most prominently thymine dimers. The bases in the dimers can no longer form hydrogen-bonded base pairs.
(vi) Strand breaks. Either directly or secondary to base alterations, phosphordiester bond breaks (single or double) follow ionizing radiation or action of certain chemical agents (e.g. bleomycin).
(vii) Crosslinkage of strands. Bifunctional alkylating agents, mitomycin C, and photoactivated psoralen adducts sometimes form covalent bridges between strands preventing their separation. (Kursiv und unterstrichen von Kornberg und Alberts et al.)
- Und er fügt hinzu (pp. 608/609):
Although the lesions and mutations produced by some agents can be predicted, in most instances the result is conditioned by the chemistry of the cell at the time the agent is acting and by a variety of replication and repair responses the cell may make to the injury. In rare instances, the lesion may so alter function as to provide selective advantage for evolution.
Den letzteren Punkt müssen wir uns noch genauer ansehen, denn auf der Allelsubstitution beruht nach wie vor ein großer Teil aller heutigen Erklärungen zum Ursprung der Arten. So schreiben auch Alberts et al. (1983, p. 106):
Very rarely, a mistake in base-pairing will lead to an improved gene. It may change a protein so that it becomes a "better" enzyme or contributes to a more effective structure. In these rare cases, organisms carrying the mutation will have an advantage, and the mutated gene may eventually replace the original gene in most of the population through natural selection.
- Und auf der Seite 771 stellen die Autoren fest:
Evolution depends to a large extent on mutations that alter existing genes to create in their place new alleles, or variants, of these genes. (Kursiv von Kornberg und Alberts et al.)
(Ähnlich Gottschalk 1984, Vogel und Angermann 1984, Günther 1984 u.v.a..) Simmonds schreibt (1979, p. 14): "The essential feature of Neo-Darwinian micro-evolution...emerges as gene substitution". Er hebt in diesem Zusammenhang jedoch die Polyploidie als Voraussetzung für Gensubstitutionen hervor, worauf wir beim Thema Duplikationen zurückkommen möchten.