3) Zeigt der Ursprung der Vogelfeder nicht deutlich, dass Gregor Mendel sich doch geirrt hat?
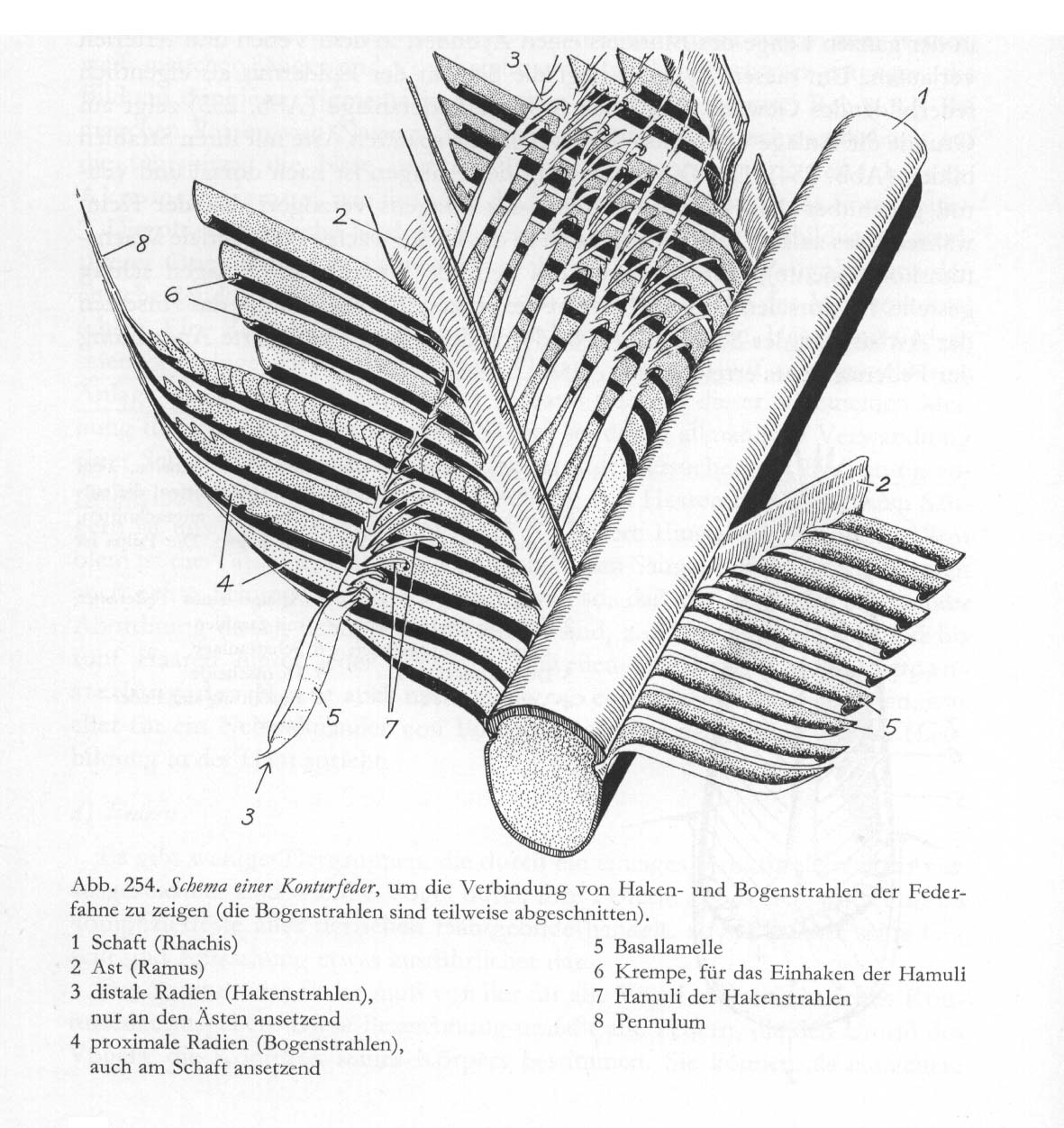
Abb. 3
N.: Abbildung aus A. Portmann (1969, S. 308): Einführung in die vergleichende Morphologie der Wirbeltiere; Schwabe & Co. Verlag, Basel und Stuttgart. Originalvorlage wesentlich klarer und deutlicher als ich Sie hier zur Zeit wiedergeben kann.
Wird am Beispiel des Ursprungs der Vogelfeder nicht letztlich doch deutlich, dass Mendels Schlussfolgerung von der genetischen "Bestätigung" der Gärtnerschen Auffassung, "dass der Species feste Grenzen gesteckt sind, über welche hinaus sie sich nicht zu ändern vermag" keineswegs korrekt war? Zu dieser Frage gebe ich meine ausführliche Diskussion mit Herr A. wieder.
[N.: Trotz aller intensiven Bemühungen um Genauigkeit und Richtigkeit meiner Ausführungen ist mir dabei ein Fehler jener Kategorie unterlaufen (vgl. Nachtrag), von dem man hinterher nur sagen kann, dass er doch eigentlich vermeidbar gewesen wäre. Dadurch ist jedoch ein unfreiwilliges, wenn auch sehr aufschlussreiches Experiment entstanden: Wäre wenigstens ein Teil der von Herrn A. beanspruchten besseren Kenntnisse und absoluten Sicherheit zum Thema durch Tatsachen abgesichert gewesen, dann hätte er meinen Fehler sofort bemerkt. Er hätte ihn in seiner Replik als wichtiges Gegenargument zur Proteinstruktur der Vogelfeder aufführen und somit die Richtigkeit zumindest dieses einen Punktes seiner Ausführungen unterstreichen können (auch wenn das an der Widerlegung seiner evolutionären Deutung der Federherkunft im Prinzip wenig oder nichts geändert hätte). Er hat den Fehler jedoch weder aufgeführt noch - soweit aus seiner Reaktion zu schließen ist - überhaupt bemerkt.]
Mein Brief vom 2.11.99
:"In der Diskussion mit Ihnen stehe ich nach wie vor vor einem schwierigen methodischen Problem (vgl. Sie bitte dazu meinen Brief vom 12. 10. 99 und meine e-Mail vom 19.10.99): Wie kann ich mir sicher sein, dass Sie meine Arbeit damit belohnen, dass Sie sie auch gründlich studieren? Wie kann ich Sie dazu motivieren, meine sorgfältig auf Richtigkeit ausgesuchten, viele relevante biologisch-wissenschaftliche Tatsachen beschreibenden Zitate und erklärenden Ausführungen, auch sorgfältig zu lesen, zu prüfen und auf ihren Wahrheitsgehalt genau zu untersuchen? Ein (weiterer) Evolutionstheoretiker, mit dem ich dieses Problem diskutierte, meinte ironisch: "Immer nur eine Seite schicken und die in Großbuchstaben!"
Aber da wir uns wohl völlig einig sind, dass wir so keine wissenschaftliche Diskussion führen können, sende ich Ihnen das, was ich zum Ursprung der Vogelfedern zu sagen habe, als zusammenhängende Abhandlung.
Nun zum Thema Herkunft der Federn:
Die Behauptung, dass die hochkomplexen Strukturen der Federn durch Mutation und Selektion in einer kontinuierlichen Evolution aus Reptilschuppen hervorgegangen sind, wird seit Jahrzehnten in Lehr- und anderen Fachbüchern (sowie in der populärwissenschaftlichen Literatur) rund um den Erdball als unbezweifelbar "wahr", d. h. als Tatsache, gelehrt. Dabei bedient man sich häufig einer (für mich als Naturwissenschaftler) erstaunlichen Methode der Desinformation, indem man nämlich längst überholte Hypothesen als nach wie vor vollkommen zutreffend verbreitet, - wie das übrigens auch beim "Biogenetischen Grundgesetz" seit mehr als hundert Jahren schon geschieht. So liest man in dem angesehenen und vielgebrauchten Spektrum-Standardlehrbuch der BIOLOGIE 1997, p. 712: "Federn bestehen aus beta-Keratin — demselben Protein, das auch die Schuppen der Reptilien bildet"[vgl. Nachtrag 2001]. Lehninger et al. geben in ihrem PRINCIPLES OF BIOCHEMISTRY (1993, p. 171) für die Vogelfeder alpha-Keratin an, welches auch bei Reptilien vorkommt. Das weiß man jedoch schon seit etwa 1972/73 besser. Die neuen Erkenntnisse der Forschung eignen sich allerdings nicht mehr so gut für die Verbreitung neodarwinistischer Hypothesen. Wie die folgende Diskussion Ihrer Aussagen zeigt, sind Sie von Evolutionstheoretikern bedauerlicherweise sachlich nicht richtig informiert worden.
Als Kommentar zu Ihren Aussagen zitiere ich im Folgenden u. a. mehrere Evolutionstheoretiker, die sich ihr Leben lang mit ornithologischen Fragen beschäftigt haben. So hat zum Beispiel Prof. Alan Howard Brush (Department of Physiology and Neurobiology, University of Connecticut, Storrs, USA) seit Anfang der 70er Jahre über die Frage der Federstrukturproteine selbst experimentell gearbeitet (er gilt zur Zeit als der weltbeste Kenner der Materie) und Prof. Adolf Portmann und Mitarbeiter haben etwa 40 Jahre (unter anderem) über Federstrukturen an seinem Institut in Basel geforscht. Desweiteren beschäftigt sich der Ornithologe Prof. Frank B. Gill (Curator and Chairman of the Department of Ornithology at the Academy of Sciences, Philadelphia) seit Ende der 60er Jahre u. a. mit der Entstehung der Federn. (Alle Hervorhebungen im Schriftbild der Zitate sind von mir.)
Zu meiner Frage an Herrn B. ("Welche naturwissenschaftlichen (hier vor allem experimentalgenetische und molekulare) Beweise gibt es für die Auffassung, dass aus Schuppen Federn entstanden sind?") führen Sie fünf Punkte auf:
Herr A.: (1) "Aus dem chemischen Aufbau der Federstruktur läßt sich die Entwicklung aus Reptilien-Schuppen jedoch zweifelsfrei herleiten."
Prof. A.H. Brush bemerkt zu dieser Frage 1996, p. 140 (On the Origin of Feathers: J. Evol. Biol 9: 131-142): "At the morphological level feathers are traditionally considered homologous with reptilian scales. However, in development, morphogenesis, gene structure, protein shape and sequence, and filament formation and structure, feathers are different" .
Mehr als diese Unterschiede der Vogelfeder im Vergleich zur Reptilschuppe auf praktisch allen Ebenen -
- kann man für eine auf Tatsachen beruhende Verneinung Ihrer obigen Behauptung kaum erwarten:
(1)"Aus dem chemischen Aufbau der Federstruktur läßt sich die Entwicklung aus Reptilien-Schuppen jedoch keineswegs zweifelsfrei herleiten. Vielmehr sind durch den chemische Aufbau die bisherigen neodarwinistischen Hypothesen widerlegt worden."
Könnte überdies in Ihrer obigen Aussage nicht vielleicht auch ein grundsätzlicher methodischer Fehler stecken? Zur Erhellung der Frage einige Analogiebeispiele: "Aus dem chemischen Aufbau der Frontscheibe des Autos läßt sich die Entwicklung aus dem Schaufensterglas jedoch zweifelsfrei herleiten." (Selbst wenn das zuträfe, dann hätte diese ‘Entwicklung‘ nicht von selbst, sondern nur unter Einsatz von Intelligenz stattgefunden.) Oder: "Aus dem chemischen Aufbau der Bücher Goethes läßt sich die Entwicklung aus der Jenaer Zeitung jedoch zweifelsfrei herleiten." Oder: "Aus dem chemischen Aufbau der Bildwerke van Goghs läßt sich die Entwicklung aus Rembrandts Werken jedoch zweifelsfrei herleiten." Reicht denn der chemische Aufbau, um etwas abstammungstheoretisch herzuleiten? Oder setzt diese Methode nicht vielmehr schon die unbewiesene Abstammungslehre als Tatsache voraus?
Herr A.: (2) "Die sekundäre Faltblattstruktur der Proteine reptiler Schuppen gleicht jener der Vogelfedern auf frappierende Weise. Gleiches trifft auf die Elementaranalyse zu."
Selbst wenn das zuträfe, wäre das kein Beweis für die Richtigkeit von Punkt (1). Denn Ähnlichkeit sekundärer Faltblattstrukturen kommt auch bei Tausenden von Proteinen vor, die man stammesgeschichtlich eindeutig nicht voneinander ableiten kann (vgl. fast jedes neuere Biochemie-Lehrbuch und Lönnig 1998). Gleiches gilt für Elementaranalysen.
Überdies widersprechen die neueren Forschungsergebnisse Ihren unter Punkt (2) aufgeführten Aussagen: "Protein sequence and shape" are different. "Amino-acid composition, shape and behavior of feather keratins are unique (einzigartig!) among vertebrates" (Brush 1996, p. 131). "There are three groups of keratins: the alpha-keratins, the beta-keratins and the feather keratins" (D. Parry 1999, p. 1343 in: Encyclopaedia of Molecular Biology, Vol. 2) [vgl. jedoch Nachtrag 2001].
"In reptiles, alpha-keratins provide the structural proteins of the skin and the flexible portions of scales (Alexander, 1970; Baden et al., 1974). A protein with an X-ray diffraction pattern similar to avian phi-keratins (Baden and Maderson, 1970) is present in the harder structures, but the structural polypeptide differs from the avian phi-keratin in size, amino acid composition, and chemical behaviour. The structure may be convergent simply as a result of functional demands" (Brush 1993, pp. 128/129).
Wohl genauer könnten Ihre obigen Sätze wie folgt lauten:
(2) [Nachtrag: Korrektur] "Abgesehen davon, dass sich die Vogelfederkeratine in der Größe des Polypeptids, der Aminosäurenzusammensetzung und des chemischen Verhaltens von denen der Reptilschuppen deutlich unterscheiden (also Merkmale aufweisen, die gegen die Abstammung der Federn von Schuppen sprechen), bestehen die Reptilschuppen und die Vogelfedern aus beta-Keratinen. Für Abstammungsfragen ist der letztere Punkt jedoch irrelevant, da es zahlreiche Beispiele für nicht voneinander abstammende Proteine mit gleicher Faltblattstruktur gibt. Eine genaue Elementaranalyse wird bei stark unterschiedlichen Proteinen auch zu unterschiedlichen Ergebnissen führen."(I)
Herr A.
(3) "Ein Mechanismus, der aus der "Zerschlitzung" der schuppenartigen Reptilienhaut zum Federkleid der Vögel führte, wurde von B(r)ush (1993) detailliert beschrieben (1)."Ich muss gestehen, dass ich immer wieder überrascht bin, mit welchem vollkommenen Sicherheitsanspruch auf absolute Wahrhaftigkeit Sie zweifelhafte evolutionistische Hypothesen zitieren. An einer detaillierten Beschreibung eines Mechanismus, der aus der "Zerschlitzung" der schuppenartigen Reptilienhaut zum Federkleid führte" (Indikativ!) könnte es nämlich — die Richtigkeit der Behauptung vorausgesetzt — auf naturwissenschaftlicher Ebene keinen Zweifel mehr geben. Und genau dieser Eindruck wird weltweit in der evolutionistischen Literatur auch zu den verschiedensten Fragen erweckt. In Wirklichkeit handelt es sich jedoch um "Glaubensaussagen". Wäre es nicht der Wahrheitsfindung dienlicher, diesen Glauben, dieses absolute "Für-wahr-Halten" unbewiesener Hypothesen, wissenschaftlich-kritisch zu hinterfragen - statt diesen Glauben völlig unkritisch und unreflektiert wiederzugeben?
In der von Ihnen erwähnten Arbeit von Brush 1993 beschreibt der Autor nun keineswegs "einen Mechanismus, der aus der "Zerschlitzung" der schuppenartigen Reptilienhaut zum Federkleid führte". Vielmehr hält er solche Hypothesen für unzulänglich. Der Titel seines Beitrags lautet entsprechend: The Origin of Feathers: A Novel Approach (Chapter 2 in: D. S. Farner et al.: Avian Biology, Vol IX, pp. 121-162, 1993). So bemerkt Brush in diesem Beitrag, pp. 124, 127, 128 u.a.:
"
Traditional hypotheses (Parkes, 1966; Maderson, 1972a,b; Regal, 1975; Dyck, 1985) deal with potential morphological intermediates and the selective forces that drove their evolution. In each case there is a serious problem of cause and effect, which is complicated by the paucity of fossil evidence.""It is not my intention to attempt to choose among the possible hypotheses of feather evolution or to suggest an alternative "cause"." Und: "If the best morphological models of the origin of feathers from scales are unsupported, it is appropriate to investigate other systems: Evidence from other systems might suggest alternative hypotheses."-
Zu den Mechanismen der ‘Zerschlitzungs-Hypothesen‘ bemerkt Brush u.a. (p. 124)
:"…there have been no convincing demonstrations of mechanisms for the conversion of a scale to a feather…Ewart (1921) was probably the first to dispute the traditional scale-feather relationship."
Im Gegensatz zur neodarwinistischen Idee der über Jahrmillionen verlaufenden kontinuierlichen Evolution von der Reptilschuppe zur Vogelfeder, präsentiert Brush nun Hypothesen, die
("Indeed it is not clear if feathers arose directly from a reptilian scale, secondarily through an avian scale, or if they shared a common ancestor" - p. 127)
und
Brush arbeitet also mit Vorstellungen, von denen B. behauptete: "Nach Makromutationen, die für die sogenannte Makroevolution oder transspezifische Evolution verantwortlich gemacht werden sollen, wird in der Forschung schon lange nicht mehr gesucht" — vgl. Sie bitte dazu meinen Brief vom 14./15. 11. 1998. (Zu Brushs Hypothesen siehe die vorliegenden Ausführungen pp.5/6)
Brush stellt jedoch zur Schlüsselfrage des Ursprungs der Federkeratingene fest (p. 131): "The origin of the phi-keratin gene family is unknown". Und nach einigen weiteren Überlegungen und Hypothesen (p.132): "There is no direct evidence from amino acid sequence that phi- and alpha-keratins diverged from a common ancestral gene. Nor is there evidence that the phi-keratins were derived from existing alpha-keratin genes."
Auf Seite 152 faßt Brush diesen Hauptpunkt wie folgt zusammen:
"The genes that direct synthesis of the avian phi-keratins represent a significant divergence from those of their reptilian ancestor. The exact nature of their origin is still unknown but they are a derived character of birds."
Tatsache ist, dass auch die anderen Evolutionstheoretiker mit ihren traditionellen ‘Zerschlitzungs-Hypothesen‘ keinen Mechanismus beschreiben, sondern nur (einander widersprechende) funktionale Hypothesen aufstellen, die in zahlreichen Punkten anfechtbar sind (vgl. Brush oben). Hier liegt also ein großer Unterschied zu einem als sicher erkannten und beschriebenen 'Mechanismus' vor! Es ist naturwissenschaftlich nicht zulässig, eine funktionale Hypothese mit einer Tatsachenbeschreibung gleichzusetzen: "Such functional hypotheses remain completely speculative. We know virtually nothing about the behaviour and ecology of the reptiles that evolved into birds" (F.B.Gill: Ornithology; W.H.Freeman & Co New York; 1990, p. 33). Richtiger könnte Ihr Satz lauten:
(3)"Sich widersprechende Hypothesen eines prinzipiell nicht reproduzierbaren "Mechanismus", der aus der "Zerschlitzung" der schuppenartigen Reptilienhaut zum Federkleid der Vögel geführt haben soll, wurde von mehreren Evolutionstheoretikern beschrieben. Brush betrachtet solche Hypothesen als unzureichend und versucht das Problem mit einer ‘non-Darwinian‘ genetischen und epigenetischen Revolution (unter ‘rapid phenotypic changes‘) zu lösen, ohne dabei notwendigerweise den Anschluß an ‘die Reptilschuppe‘ zu suchen. Er gesteht aber ein: "The origin of the phi-keratin gene family is unknown."
Naturwissenschaftliche Beweise für die Richtigkeit der behaupteten Evolution der Vogelfeder aus Reptilschuppen durch Mutation und Selektion fehlen völlig.
Herr A.: (4)"Eine derartige Homologie auf molekularer Ebene lässt darüber hinaus Rückschlüsse auf die Übereinstimmung in der Gensequenzierung zu, die den Aufbau der Schuppenhaut bzw. der Vogelfedern, codiert…"
(Gensequenzierung ist die Methode. Sie meinen wahrscheinlich ‘Übereinstimmung in der Nucleotidsequenz‘.)
Die naturwissenschaftlichen Tatsachen widersprechen Ihrer Aussage. Prof. F. B. Gill stellt in seiner Ornithology (W.H.Freeman & Co.; New York) 1990, pp. 57/58 fest:
"Although keratin in the generic sense is a standard constituent of other hard epidermal structures, such as claws, hair, fingernails, and scales, the phi-keratin in bird feathers is unique. It is not, as was once believed, the same as that found in reptile scales. The genes that control the production of avian phi-keratin have been isolated…and were found to be radically different from genes that control other keratin synthesis (Gregg et al. 1983, 1984). It appears that a genetic revolution of some kind led to the novel chemistry of feathers (Brush 1989)."
Und Brush selbst bemerkt 1996, p. 132 und p. 136:
The molecular evidence questions the simple, direct relation of the specialized structures of birds to reptile scale. I will provide arguments to show that reptilian scales and feathers are related only by the fact that their origin is in epidermal tissue. Every feature from gene structure and organization, to development, morphogenesis and tissue organization is different." - "There are no compelling matches between avian feather, scute, claw or beak and proteins from reptilian epidermal structures…"
Herr A.
: (5) "…(siehe auch meine spätere Ausführung zur Entwicklung des Auges auf Seite 6)."Beim Pax-6-Gen des Auges liegen die Verhältnisse anders als bei den Keratingenen.
Wie stellt sich nun der Evolutionstheoretiker A.H.Brush die Entstehung der Feder vor? Dazu seine folgenden Aussagen (alle Hervorhebungen im Schriftbild wieder von mir):
A.H. Brush (1996, p. 131, abstract): "The changes were hierarchical in nature and included revolutions in genome organization (i.e., HOX and the feather keratin genes), protein sequence and shape, the large scale organization of proteins into filaments, and in the geometry of the cells and their roles in the follicle. Changes in each of these levels differ or produced different products than found in its analog in reptiles. They are essentially unique to birds and produced an evolutionary novelty. I used analysis of extant structure and information on the development to reconstruct key events in the evolution of feathers. The ancestral reptilian epidermal structure, while probably a scale or tubercles, is still unidentified. The structural genes of feathers proteins, (phi-keratin) are tandem repeats probably assembled from pre-existing exons. They are unlike the alpha-keratin of vertebrate soft epidermis. Amino-acid composition, shape, and behavior of feather keratins are unique among vertebrates. The 3-dimensional organization of the follicle and the developmental processes are also unique. Although we lack a complete understanding of the appearance and early role of feathers, they are clearly the results of novel events." (On the Origin of Feathers. J. Evol. Biol. 9, 131-142, 1996).
"It is clear that the appearance of novel structures requires simultaneous and important changes at the genic, cellular, tissue and organismal levels (Ashley and Hall, 1991). Defining the particular changes and recognizing the emergent consequences is not always possible. The morphological revolution that produced feathers is inextricably associated with a set of closely coordinated structural genes and a unique production machine. The mechanism appears to retain linearity, but several epigenetic processes are involved" (Brush, p. 140).
"Clearly, feathers provide a unique and outstanding example of an evolutionary novelty" (Brush, p. 140).
Das klingt nun nicht mehr nach neodarwinistisch-kontinuierlicher Evolution: Statt dessen postuliert der Autor Revolutionen auf allen Ebenen: (1) Revolution in der Genom-Organisation mit HOX- und den Federkeratin-Genen, (2) Revolution in der Proteinsequenz und -form, (3) Revolution der großangelegten Organisation der Proteine zu Filamenten, und (4) Revolution in der Geometrie der Zellen und ihrer Rolle in den Follikeln.
Er räumt dabei ein, dass die Erklärung und Definition der besonderen Veränderungen und ihrer "emergent consequences" nicht immer möglich ist. Auf der DNA-Ebene greift der Autor vor allem auf die Exon-Shuffling-Hypothese zurück, die er jedoch nicht hinreichend beweisen kann — genausowenig wie die von ihm postulierten evolutionär-epigenetischen Prozesse.
Die neodarwinistische Idee von sich in Millionen von Jahren kontinuierlich anreichernden Punktmutationen mit korrelierter morphologischer Divergenz lehnt der Autor jedenfalls klar ab. Er postuliert vielmehr "gene rearrangements" und deren "rapid incorporation into the genome" als eine Folge von "molecular drive". "The behavior of the genes is independent of population size or local selective regimes." ‘Radikale phänotypische Veränderungen‘ sollen dabei auf recht schnelle Veränderungen in der Gen- und Genomstruktur zurückzuführen sein etc..Keine dieser Hypothesen läßt sich für die Federenstehung beweisen.
Kommen wir zu unserem Ausgangspunkt zurück (Frage an Herrn B.): "Welche naturwissenschaftlichen (hier vor allem experimentalgenetische und molekulare) Beweise gibt es für die Auffassung, dass aus Schuppen Federn entstanden sind?"
Zusammenfassend ist festzustellen, dass die biologischen Forschungsergebnisse allen fünf(Korrektur: "Nur" vier [oder viereinhalb]: die Faltblattstruktur ist richtig, aber irrelevant, vgl. Vorwort.) der von Ihnen zitierten Punkte widersprechen.
Es gibt keinerlei empirische Evidenz, dass durch selektierte Zufallsmutationen die komplexen Strukturen der Vogelfedern entstanden sind. Die molekularen und morphologischen Befunde zeigen vielmehr, dass Federn in der
einzigartig sind und sich somit in fast jeder Hinsicht deutlich von Reptilschuppen unterscheiden.
Der Ursprung der Vogelfeder wird weder durch den Neodarwinismus noch durch andere bekannte Evolutionstheorien naturwissenschaftlich hinreichend erklärt .
Zur Frage nach einer Alternative zur Evolutionstheorie:
Sie werden mich an dieser Stelle wahrscheinlich fragen, wie ich denn den Ursprung der Vogelfeder erkläre — ganz im Sinne Ihrer letzten e-Mail, aus welcher ich im Folgenden zitiere:
Herr A. (e-Mail vom 25. Oktober 1999): "Ehrlich gesagt, ich bin mir immer noch nicht im klaren darüber, welche Theorie Sie nun eigentlich vertreten, um die Phylogenese wissenschaftlich zu beschreiben."
Setzen Sie nicht in dieser Anfrage die unbewiesene "Phylogenese" als Tatsache bereits voraus und machen damit eine Glaubensaussage? Vgl. Sie bitte meinen Brief an Herrn Q. vom 9. 12. 99, pp. 10/11, wo zahlreiche Evolutionstheoretiker zugeben, dass "die Phylogenese" nicht nachweisbar ist. Eine nicht nachweisbare Phylogenese kann man auch nicht naturwissenschaftlich beschreiben.
Herr A.: "Zwar lassen Sie immer wieder mal durchblicken, dass Sie sich zu den Kreationisten rechnen ("So, why not creation?"), scheinen diesen Standpunkt aber nicht offensiv zu vertreten, sondern formulieren nur beständig den Unmöglichkeitscharakter der Evolutionstheorie."
Ich rechne mich nicht zu den Kreationisten. Zur Definition des Kreationismus gehört nach Science vom 22. Oktober 1999, p. 659 (und ebenso nach Auffassung der Kreationisten selbst):"…creationists believe that God created the universe in 6 days 10,000 years ago…"
(Vgl. Sie dagegen bitte meinen Brief an Herrn Q. vom 14./15.11. 98, p. 20: "Im übrigen sollten wir unsere Gesprächspartner einmal davon in Kenntnis setzen, dass wir, d. h. Sie und ich, keine Kreationisten sind. Wir glauben weder an buchstäbliche 24-Stunden-Schöpfungstage, noch dass das Universum nur 10 000 Jahre alt ist, noch an die Dreieinigkeit, noch an die Hölle (in welche nach Auffassung der Kreationisten die Evolutionisten u. a. kommen) etc.").
Wir haben also im Vergleich zu den Kreationisten, die sich zumeist an konservativen und kirchlichen Dogmen orientieren, in zahlreichen und grundlegenden Punkten ein völlig abweichendes Bibelverständnis. Darüber hinaus versuchen verschiedene kreationistische Gruppen, ihre Kritik mit politischen Mitteln durchzuzetzen — wir grundsätzlich nicht (was den Griff nach politischen Instrumenten betrifft, stehen übrigens in den USA die Evolutionisten den Kreationisten keineswegs nach). Wenn Sie jedoch alle Personen, die die Bibel ernstnehmen und an eine Schöpfung für wahr halten ("So, why not creation?"), zu den Kreationisten rechnen, dann werden Sie uns sicherlich dazuzählen. Allerdings halte ich diese Einstufung für so ungenau wie wenn jemand (die) Sozialdemokraten zu den Kommunisten rechnet oder (die) Christdemokraten rechtsradikalen Gruppen zuordnet. Vom dogmatischen Kreationismus unterscheidet sich mein Schöpfungsverständnis auf biologischer Ebene insbesondere darin, dass es nicht obligat, sondern fakultativ ist: Es orientiert sich an den biologischen Realitäten (vgl. Brief vom 9.12.98, pp. 19/20). In Anlehnung an die Begriffe Evolutionstheoretiker und Evolutionsbiologe nenne ich mich und Gleichgesinnte Schöpfungstheoretiker und Schöpfungsbiologen. (Evolutionismus und Kreationismus bezeichnen die Dogmatiker.)
[Korrekturen: Nachtrag Mai 2002: Die rein wissenschaftliche Bezeichnung ist "Intelligent-Design-Theoretiker". Nachtrag September 2002: In der Internet-Version der Augenschrift wurden zu dieser Frage in der Vorbemerkung von 2002 die folgenden weiteren Punkte hervorgehoben: "Eine Korrektur habe ich in der Internetausgabe der Schrift mehrmals vorgenommen: Statt von Schöpfungslehre spreche ich in der Regel von der Intelligent-Design-Theorie, um damit die wissenschaftliche Alternative zur Synthetischen Evolutionstheorie in dieser Arbeit über die Entstehung des Auges zu betonen. Denn Intelligent Design ist auch ohne Religion wissenschaftlich fassbar und nachvollziehbar (d.h. "...it does not depend on religious premisses" - M. Behe 2002; vgl. weiter William A. Dembski (2002): NO FREE LUNCH; Roman and Littlefield, Lanham). Selbstverständlich stellt sich nach Erweis von Intelligent Design die Frage nach dem Designer. Die Fragen, ob jemand den Designer (1) mit Gott und weiter (2) mit dem Gott der Bibel identifiziert (beides ist desöfteren aufgrund zahlreicher weiterer Evidenzen der Fall) - sind rein logisch jedoch zunächst einmal unabhängig von der Entdeckung einer plan- und sinnvollen Entstehung der Organismen. Einige Forscher lassen diese Fragen offen. Michael Behe hat dazu ein aufschlussreiches Kapitel in seinem Buch DARWINS BLACK BOX (1996) geschrieben (das Zitat oben stammt aus einem Interview aus dem Video UNLOCKING THE MYSTERIES OF LIFE, Illustra Media, 2002)." - Ich spreche also auf der rein naturwissenschaftlichen Ebene statt von Schöpfungstheoretikern bzw. Schöpfungsbiologen von Intelligent-Design-Theoretikern und Intelligent-Design-Biologen. Nachtrag Oktober 2002: Wir pflegen in den Naturwissenschaften zwischen (a) der persönlichen Motivation und (b) der naturwissenschaftlichen Ebene zu unterscheiden. Aus heutiger naturwissenschaftlicher Sicht gehören (a) zu meiner persönlichen Motivation Bibel und Schöpfung und (b) zur (mathematisch-) naturwissenschaftlichen Ebene (1) die naturwissenschaftliche Argumentation und Beweisführung gegen die Synthetische (und andere) Evolutionstheorie(n) und (2) die rational-empiriokritische Argumentation für Intelligent Design (der zweite Punkt wird jedoch von vielen Neodarwinisten aus philosophischen Gründen [siehe unten] noch bestritten). - Zurück zum Text von 1999:]
Die beständige Formulierung des "Unmöglichkeitscharakters" (besser: der Grenzen) der Evolutionstheorie ist zunächst einmal notwendig, um Raum für die mit intelligenten Ursachen arbeitende Alternative zu schaffen. Desweiteren ist in einer Situation, in der es als Alternativen letztlich nur zwei Möglichkeiten gibt, jedes stichhaltige Argument gegen die eine Möglichkeit zugleich eine Bestätigung der anderen. Hier könnten Sie jedoch einwenden, dass eventuell noch eine weitere Möglichkeit besteht (siehe Portmann und von Frisch, p. 11/12 des vorliegenden Schreibens).
Herr A.: Deshalb habe ich die Bitte, mir eine kurze e-Mail zu senden, in der Sie Ihre Sicht der Dinge bezüglich der Entstehung und Entwicklung der Artenvielfalt explizit offenlegen. Vielen Dank!"
Habe ich Ihnen meine Sicht der Dinge bezüglich der Entstehung und Entwicklung der Artenvielfalt in den Ihnen übersandten Arbeiten nicht bereits mehrmals offensiv und explizit offengelegt? — Ich bin, offen gestanden, etwas erstaunt über diese Anfrage. Könnte das mit dem in der Einleitung des vorliegenden Briefes erwähnten Kommunikationsproblem zusammenhängen? Es ist unzutreffend, dass ich "nur" beständig den "Unmöglichkeitscharakter" der Evolutionstheorie formuliere.
Ich wiederhole im Folgenden ein Hauptargument meiner Aussagen, welches ich in der Diskussion mit B. und anderen aufgeführt habe.
Der Zellbiologe Siegfried Strugger bemerkte einmal treffend:
"Die Zelle ist das vollendetste kybernetische System auf der Erde. Alle Automation der menschlichen Technik ist gegen die Zelle nur ein primitives Beginnen des Menschen, im Prinzip zu einer Biotechnik zu gelangen."
Wenn nun schon "das primitive Beginnen" auf diesem Weg immer bewußtes Handeln, Intelligenz, Geist und Weisheit voraussetzt, - wieviel mehr muß das dann auf den Ursprung der tausendmal komplexeren kybernetischen Systeme der Lebensformen zutreffen!
(Vgl. Sie bitte desweiteren meine Augenschrift, pp. 34-39, wo ich - wiederum ganz offensiv und explizit — und ganz im eben zitierten Sinne, zahlreiche weitere Details offengelegt habe, und noch ausführlicher in der Artbegriffsarbeit pp. 283-290.)
Es gibt eine reiche empirische Evidenz, dass durch bewußtes Handeln, Intelligenz, Geist und Weisheit hochkomplexe Informationen erzeugt und vielfach vernetzte kybernetische Strukturen erschaffen werden können. Kann es an der Richtigkeit dieser Aussage irgendeinen Zweifel geben?
Welche Ursachen sind dann für im Prinzip vergleichbare, aber 1000mal komplexere, Systeme zu erwarten?
Auf den Ursprung der Vogelfeder angewandt, schließe ich gleichsam (ich betone gleichsam:) auf einen "Super-Geningenieur", der (genialer als alle Gentechnologen der Welt zusammen) die Revolutionen - von der DNA (HOX- und Federkeratin-Gene) bis zur Geometrie der Zellen - verursachte. Ich nenne das die "intelligente DNA- und Zellprogrammierung" — ganz im Sinne der Genesis 1 : 21: "Und Gott ging daran…jedes gefügelte fliegende Geschöpf nach seiner Art" zu erschaffen. Statt von einer "evolutionary novelty" (siehe Brush oben, p. 6) spreche ich von der Feder als von einer "creationary novelty". (Zu weiteren Fragen, wie der nach der Bedeutung der Struktur-Ähnlichkeiten auf den verschiedenen biologischen Ebenen, vgl. Sie bitte die Ausführungen von ReMine 1993, die ich für Sie kopiert habe).
Ich möchte an dieser Stelle noch einmal betonen, dass dieser Ansatz — ganz im Gegensatz zu den verschiedenen evolutionistischen Vorstellungen zu selektierten Zufallsmutationen — eindeutig an der Erfahrung, und zwar an einer reichen empirischen Evidenz, orientiert ist.
Sie werden wahrscheinlich dennoch einwenden, dass diese Auffassung keine naturwissenschaftliche Hypothese sei. Und Sie haben auch völlig Recht mit diesem Einwand, wenn Sie naturwissenschaftlich mit naturalistisch gleichsetzen. Wenn man also sagt, dass Naturwissenschaft ausschließlich mit den uns bekannten physikochemischen Gesetzmäßigkeiten rechnen darf, dann ist die soeben vorgetragene Auffassung nicht naturwissenschaftlich — unabhängig von der Frage, ob die Antwort falsch oder letztlich doch wahr ist. Scott C. Todd schreibt in diesem Sinne (Nature 401, p. 419, Ausgabe vom 30. September 1999):
"Even if all the data point to an intelligent designer, such a hypothesis is excluded from science because it is not naturalistic. Of course the scientist, as an individual, is free to embrace a reality that transcends naturalism".
Genau genommen wird hier die naturalistische Philosophie über die Tatsachen (die Daten) gestellt. Selbst wenn alle Daten auf einen Intelligent Designer hinweisen, ist dennoch diese Schlußfolgerung in der Naturwissenschaft nicht erlaubt! (Das heißt also:…‘umso schlimmer für die Tatsachen!‘) - Das darf man wohl als Metaphysik bezeichnen!
Die Frage erhebt sich jedoch in diesem Zusammenhang, ob man die Biologie dogmatisch auf eine Methode und Philosophie beschränken muß, die nicht in der Lage ist, die volle Realität in ihren Ursachen und Abläufen zu beschreiben. Muß man den Teil der Naturwissenschaft, der sich vor allem mit Ursprungsfragen, also historischen Fragen, beschäftigt, ebenfalls auf den Naturalismus beschränken oder kommt mit der historischen Fragestellung nach den Ursprüngen nicht doch eine neue Dimension ins Spiel, welche über den physikochemischen Reduktionismus hinausgeht? Könnte die historisch nach dem Ursprung der Lebensformen fragende und arbeitende Biologie nicht auch Geisteswissenschaft sein? Die Verabsolutierung der naturalistischen Methode kann jedenfalls zu erheblichen Erkenntnisschwierigkeiten führen - wie das Beispiel aus meiner Diskussion mit Herrn Prof. D. deutlich zeigt (ich gebe es im Folgenden noch einmal wieder, vgl. Sie bitte meinen Brief vom 9.12.98, p.5).
(Prof. D.:) "Die Frage nach dem Konstrukteur ist daher tatsächlich vom Naturwissenschaftler grundsätzlich abzulehnen."
Ich stelle mir zur Veranschaulichung nur einen solchen Naturwissenschaftler vor, der - sagen wir - auf dem Mars ein ihm bisher unbekanntes komplexes und vielfach vernetztes Computer- und Datenverarbeitungssystem entdecken würde, ohne dessen Erfinder zu kennen oder ohne weiteres identifizieren zu können. Was macht er nun mit seiner naturwissenschaftlichen Denk- und Arbeitsweise? Antwort: "Die Frage nach dem Konstrukteur ist daher tatsächlich vom Naturwissenschaftler grundsätzlich abzulehnen!" Er wird folglich alle qualitativen Phänomene auf quantitative reduzieren wollen und dann eine entsprechende naturwissenschaftliche Theorie formulieren (siehe weitere Beispiele und Diskussion, Artbegriff p. 285 ff.). Und ich behaupte nun mit allem Nachdruck, dass kein halbwegs normaler Naturwissenschaftler in dieser Situation so vorgehen würde. Fast jeder Naturwissenschaftler wird nach dem intelligenten Ursprung und nach dem (oder den) Konstrukteur(en) fragen. Und genau deswegen zeige ich auch immer wieder die Identität kybernetischer Systeme in Biologie und Technik auf. Die Frage ist auf beiden Gebieten nicht nur legitim, sondern zur Wahrheitsfindung absolut unerlässlich (vgl. weiter Artbegriff S. 286).
Wenn man nicht den Naturwissenschaftler, sondern nur die Naturwissenschaft in der oben zitierten Weise beschränkt (was ja auch für weite Bereiche völlig legitim ist) und in diesem Rahmen die Schlussfolgerung von der Konstruktion auf den Konstrukteur grundsätzlich verbietet, dann kann man - um nicht völlig an den Realitäten vorbei zu gehen - auf der anderen Seite aber auch argumentieren, dass die Biologie eben mehr ist als nur beschränkte Naturwissenschaft (von Buddenbrock, Portmann, Troll, Eckardt, S.Vogel, O.Kuhn, Spemann, von Frisch, Eccles und viele andere). Die Biologie ist dann zumindest in den Ursprungsfragen auch 'Geisteswissenschaft' und damit sind diese Fragen und Schlussfolgerungen auch innerhalb der Biologie wieder legitim. Weiter darf man darauf hinweisen, dass die Schlussfolgerung von der Konstruktion auf den Konstrukteur aufgrund aller Erfahrungen sowohl logisch als auch intersubjektiv nachvollziehbar ist (vgl. weiter Artbegriff S. 288 oben).
Auf meinen Hinweis auf das evolutionistische Denkverbot (Artbegriff, p. 8: "Frage auch bei den komplexesten und genialsten Konstruktionen niemals nach dem Konstrukteur") reagierte Herr D. recht emotional:
(Prof. D.:) "Das ist kein feiges oder illegitimes oder vorurteilsbelastetes Denkverbot, sondern im Gegenteil eine Notwendigkeit für den methodisch sauber und ehrlich arbeitenden Wissenschaftler."
Welch Adjektive! (Sie stammen nicht von mir.) Da steht unser Naturwissenschaftler in der obigen Veranschaulichung vor dem komplexen und vielfach vernetzten Computer- und Datenverarbeitungssystem und will dessen Herkunft "naturwissenschaftlich", d. h. mit den zitierten Einschränkungen, erklären. Er lehnt daher die Frage nach dem Konstrukteur kategorisch ab und erklärt, das sei "kein feiges oder illegitimes oder vorurteilsbelastetes Denkverbot, sondern im Gegenteil eine Notwendigkeit für den methodisch sauber und ehrlich arbeitenden Wissenschaftler". Mehr noch, er versucht diejenigen, die die richtige Frage stellen, in ihrer Arbeit und ihrem Fortschritt zu behindern und behauptet, nur seine Fragestellung und Methodik sei sauber und ehrlich. Gleichzeitig baut er jedoch zur Ursprungsfrage ein nicht verifizierbares, nicht falsifizierbares und nicht quantifizierbares Erklärungssystem auf, in dem "der Zufall" eine entscheidende Rolle spielt. Ich meine, die Frage muss erlaubt sein, ob das wirklich noch sauber und ehrlich ist, von der Toleranzfrage ganz zu schweigen. - Was auf die Methodik zur Entschlüsselung der Funktion der Systeme zutrifft, kann bei der Frage nach dem Ursprung der Systeme völlig verfehlt sein.
(Wurde nicht auch in diesen Passagen meine grundsätzliche Sicht der Dinge bezüglich der Entstehung und Entwicklung der Artenvielfalt offensiv und explizit offengelegt? Ist eine Steigerung in der Deutlichkeit überhaupt noch möglich?)
Mir ist natürlich klar, dass zur Methodenproblematik noch sehr viel mehr zu sagen ist. (Ockhams Razor z.B. ist nur dann anzuwenden, wenn es um gleichwertige adäquate Lösungsversuche geht — niemand wird eine falsche aber einfache Lösung einer komplexeren aber wahren vorziehen.)
Zurück zur Entstehung der Feder: Könnten Sie sich nach den auf den Seiten 1 bis 7 aufgeführten biologischen Tatsachen der Meinung anschließen, dass die neodarwinistischen Voraussetzungen und Behauptungen zur Entstehung der Feder durch die neuen molekulargenetischen, biochemischen und zellbiologischen Befunde klar widerlegt sind?
Und dürfte diese Aussage nicht auch dann gelten, wenn man bislang keine Alternative zum Neodarwinismus in Betracht ziehen möchte? Denn wird ein vernünftiger Mensch argumentieren, dass er bei einer nachweislich falschen Erklärung solange bleibt, bis er eine bessere findet (-"bessere" im reduktionistischen Sinne)? Kann sich eine Hypothese nicht auch unabhängig von einer solchen Alternative als falsch erweisen? Wir kennen in den Naturwissenschaften in allen Disziplinen zahlreiche offene Probleme (davon leben wir ja als Wissenschaftler). Wenn sich eine Hypothese zu einer Frage als falsch erweist, dann ist die Frage eben erst einmal wieder offen — bis zur nächsten Hypothese/Erklärung!
Ich möchte in diesem Zusammenhang noch einmal nach den Falsifikationskriterien für den Neodarwinismus fragen (und darin erinnern, dass eine Theorie ohne Falsifikationsmöglichkeiten keine wissenschaftliche Theorie, sondern eine metaphysische Auffassung ist! — Vgl. Sie dagegen meine Aufführung von Falsifikationskriterien für die Schöpfungslehre in der Diskussion mit [Prof. K.] und auch (Herrn X.): Brief vom 9.12.98, pp. 19-20).
Sind offenen Fragen in den Naturwissenschaften erlaubt?
Obwohl Portmann glaubte, dass Reptilien die Ahnen der Vögel gewesen seien, schreibt er zum Ursprung der Federn im Sinne eines offenen Problems (Vom Wunder des Vogellebens; Piper-Verlag; 1984, p. 17/18):
"Wie ist die Vogelfeder entstanden? Dies bleibt wohl das zentrale Problem…/…für unsere Zeit müssen wir feststellen: Das Geheimnis des erdgeschichtlichen Werdens dieses erstaunlichen Hautgebildes bleibt ungelöst.
…Schuppenbildung gehört zu den Möglichkeiten aller Landwirbeltiere und kommt darum bei Kriechtieren, Säugern und Vögeln in vielerlei Varianten vor. Sie formt verhältnismäßig einfache Gebilde. Federn und Haare dagegen sind viel kompliziertere Sondergestalten, die mit Nervensystem, Blutgefäßen, Muskeln und mit Drüsen in Beziehung stehen können. So suchen manche Forscher heute die Spuren des Ursprungs von Haaren bei den Amphibien, weil die Idee einer sehr langen besonderen Geschichte der Hautgebilde von Vögeln und Säugern sich durchsetzt. Es liegt nahe, auch die Entstehung der Federn unabhängig von Kriechtierschuppen zu verstehen, als eine ganz besondere Leistung der Haut warmblütiger Tiere."
George C.Kent und Larry Miller stimmen dem insofern zu, als sie darauf hinweisen, dass die Entstehung der Federn aus Reptilschuppen als fraglich betrachtet wird und hinzufügen (Comparative Anatomie of the Vertebrates, 8thed., W.C.Brown; 1997, p. 110:):
"The only other hypothesis that is justified at present is that, like hairs, they may be new evolutionary structures lacking a reptilian ancestor" (zit. nach mehreren Quellen).
(Mit dem Thema des Ursprungs der Haare erwähnen die Autoren - wie oben schon Portmann — gleich noch ein weiteres ungelöstes Evolutionsproblem.)
Richard Cowen weist zur Frage nach der Federentstehung 1995, p. 258 auf folgenden Schlüsselpunkt hin (History of Life. Sec. Ed., Blackwell):
"Feathers in living birds originate in a deep skin layer under the outer layer that forms scales. Evolutionarily, then, feathers probably arose under and between reptile scales, not as modified scales."
(Die Situation ist jedoch noch komplizierter, indem verschiedene Zellschichten synorganisiert an der Federbildung mitarbeiten, aber die Details möchte ich an dieser Stelle nicht diskutieren.)
Portmann schreibt in Anspielung auf Darwins Äußerung, dass ihn ‘heute eher kleine, aber irritierende Eigenschaften der Struktur beunruhigen‘ und der Anblick einer Pfauenfeder krank mache, - zur Frage nach dem Ursprung der Federn und der Richtigkeit der Selektionstheorie (An den Grenzen des Wissens; Econ. 1974, S. 38):
"Seit etwa 40 Jahren sind in unserem Basler Institut diese >eher kleinen, aber irritierenden Eigenschaften der Struktur<, mit Gestalt und Farbe der Vogelfeder, ein wichtiges Objekt unserer Arbeit. Daunenkleid und Schillerfarben, Jugend- und Alterskleid, die Schmuckfeder der Mandarinente und vieles andere ist untersucht worden. Vor allem aber sind drei umfassende Studien über das Pfauenrad und zwei über die Prachtfedern des männlichen Argusfasans in Basel entstanden. Sie haben viele neue Aufschlüsse gebracht, aber das Rätsel haben sie nicht gelöst. Im Gegenteil: Es ist immer unwahrscheinlicher geworden, dass der Selektionsvorgang, von dem Darwin ausging, ein entscheidender Faktor beim Formwerden, bei der Entstehung dieser seltsamen, wunderbaren Gebilde gewesen ist. Wir haben uns konzentriert auf das Geschehen im Federkeim, der das strahlende Pfauenauge oder die Kette der Augenflecke auf den Flügelfedern beim Argushahn hervorbringt. Das Studium des Entstehens der konzentrischen Schillerbildungen beim Pfau oder der Erzeugung beim Argusfasan — beides weist auf komplizierte Strukturveränderungen in der frühen Anlage des Federkeims hin, durch welche so komplexe optische Wirkungen erreicht werden. Eine Mitwirkung der anschauenden Vogelaugen ist wohl für die Erhaltung der bereits entwickelten Struktur möglich, an den verborgenen Anlässen, die im Keim zu diesem Ergebnis führen, haben aber diese Augen keinen Anteil. Die Wirkung der fertigen Struktur auf die Henne hat diese Erscheinung >zum Anschauen< nicht geschaffen."
Portmann betont an zahlreichen Stellen seiner Werke, dass die uns bekannten richtungslosen Mutationen (plus Selektion) die Entstehung der Lebensformen nicht erklären können. Er ließ die Frage nach den Ursachen des Ursprungs der Lebensformen offen und bemerkte an einer Stelle: "Ich weiß, wie besonders schwer diese Zurückhaltung ist, wo man doch endlich einmal über die Frage unseres Ursprungs klaren Bescheid haben möchte" (1984, p. 107). Aber eine offene Frage war ihm lieber als eine falsche Antwort. (Übrigens hätte auch ein Hoimar von Ditfurth diese Studien Portmanns und seiner Arbeitsgruppe kennen und beurteilen können.)
Ein weiterer großer Biologe des 20. Jahrhunderts, der Nobelpreisträger Karl von Frisch, fasste seine Forschungserfahrung zur Ursprungsfrage in einem Fernsehinterview wie folgt zusammen:
"Ich bin überzeugt, dass mit Mutation, wie wir sie jetzt kennen, und Selektion allein nicht entfernt die Vorgänge in der lebenden Natur erklärt werden können. Sie sind einfach zu kompliziert und verwickelt als dass das denkbar wäre."
Auch er ließ die Frage nach den Ursachen der Entstehung der Lebensformen offen.
Um eventuellen Missverständnissen vorzubeugen, möchte ich darauf hinweisen, dass die letzteren Zitate nicht etwa als "Gegenbeweis" zu Ihren Punkten (1)-(5) gedacht sind (diesen haben ja schon die biologischen Tatsachen geliefert, welche vor allem von Brush, Gill, Parry und Cowen aufgeführt wurden). Zur Ergänzung der Beweislage könnte ich jetzt noch auf die Details von Portmanns Argumentation gegen eine neodarwinistische Deutung seiner Studien der Vogelfeder zu sprechen kommen. Portmann und von Frisch haben jedenfalls an der Aufdeckung der ungeheuren Komplexität lebender Strukturen erfolgreich mitgearbeitet und im Zusammenhang mit der Ursprungsfrage und der Diskussion des Neodarwinismus gezeigt, dass man sehr wohl mit "offenen Fragen" leben und wissenschaftlich gründlich arbeiten kann.
ZUSAMMENFASSUNG/WIEDERHOLUNG DER HAUPTPUNKTE:
Beim Vergleich zwischen Reptilschuppe und Vogelfeder sind zahlreiche grundlegende Unterschiede festgestellt worden. Die Feder zeichnet sich im Vergleich zur Reptilschuppe aus durch:
unterschiedliche ontogenetische Entwicklung
unterschiedliche Morphogenese
unterschiedliche Genstruktur
unterschiedliche Proteingestalt und —sequenz
unterschiedliche Filamentbildung und —struktur
.Es gilt:
(1) "Aus dem chemischen Aufbau der Federstruktur läßt sich eine Entwicklung aus Reptilien-Schuppen jedoch keineswegs zweifelsfrei herleiten. Vielmehr sind durch den chemischen Aufbau die bisherigen neodarwinistischen Hypothesen widerlegt worden."
(2)[Nachtrag: Korrektur] "Abgesehen davon, dass sich die Vogelfederkeratine in der Größe des Polypeptids, der Aminosäurenzusammensetzung und des chemischen Verhaltens von denen der Reptilschuppen deutlich unterscheiden (also Merkmale aufweisen, die gegen die Abstammung der Federn von Schuppen sprechen), bestehen die Reptilschuppen und die Vogelfedern aus beta-Keratinen. Für Abstammungsfragen ist der letztere Punkt jedoch irrelevant, da es zahlreiche Beispiele für nicht voneinander abstammende Proteine mit gleicher Faltblattstruktur gibt. Eine genaue Elementaranalyse wird bei stark unterschiedlichen Proteinen auch zu unterschiedlichen Ergebnissen führen."(I)
(3) "Sich widersprechende Hypothesen eines prinzipiell nicht reproduzierbaren "Mechanismus", der aus der "Zerschlitzung" der schuppenartigen Reptilienhaut zum Federkleid der Vögel geführt haben soll, wurde von mehreren Evolutionstheoretikern beschrieben. Brush betrachtet solche Hypothesen als unzureichend und versucht das Problem mit einer ‘non-Darwinian‘ genetischen und epigenetischen Revolution (unter ‘rapid phenotypic changes‘) zu lösen, ohne dabei notwendigerweise den Anschluß an ‘die Reptilschuppe‘ zu suchen. Er gesteht aber ein: "The origin of the phi-keratin gene family is unknown."
(4)"Die von neodarwinistischen Autoren für die Reptilschuppe und Vogelfeder fest erwartetete Homologie auf der Proteinebene mit Rückschlüssen auf Übereinstimmung in der Nucleotidsequenz ist durch die molekularbiologische Forschung widerlegt worden."
(5) "Ein Vergleich zwischen den Federkeratingenen und dem Pax-6-Gen ist verfehlt."
(6) "Aufgrund der ungeheuren Komplexität der an der Federbildung beteiligten eigenständigen molekularen und biologischen Prozesse räumen die besten Kenner der Materie (wie Brush, Gill und Portmann) ein, dass die grundlegenen Fragen zur Entstehung der Feder auch nach fast 150 Jahre währenden evolutionistischen Spekulationen nicht nur noch offen sind, sondern dass mit zunehmendem Wissen eine neodarwinistische Evolution immer unwahrscheinlicher wird."
(7)" Zahlreiche an der Erfahrung orientierte Biologen schließen auf Intelligent Design als notwendige und adäquate Ursache für den Ursprung hochkomplexer synorganisierter (d.h. sich durch vielfache und vielschichtige Koaptationen auszeichnende) biologischer Systeme, wie das der Vogelfeder."
Zur Problematik der Entstehung der Vogelfeder wäre noch sehr viel mehr zu sagen (so habe ich z. B. das Synorganisationsproblem bisher nur angedeutet), aber ich denke, dass ich mit dem vorliegenden Schreiben schon mehrere ganz entscheidende biologische Fakten aufführen und diskutieren konnte. Gerne möchte ich Sie einladen, zu den hier aufgeführten Punkten Stellung zu nehmen. Vielleicht könnten Sie mit den ersten fünf Punkten anfangen, da sie zu den direkten Antworten zu Ihrer wie folgt formulierten Anfrage vom 11. Oktober 1999 gehören: "Ich bitte Sie daher, Punkt für Punkt auf meine Argumentation einzugehen." Mit der nochmaligen Bitte um ein gründliches Studium dieses Briefes im eingangs erwähnten Sinne (die Ausführungen sorgfältig zu lesen, zu prüfen und auf ihren Wahrheitsgehalt genau zu untersuchen!) möchte ich an dieser Stelle erst einmal meinen Brief unterbrechen. Das nächste Mal möchte ich Ihren neodarwinistischen Lösungsversuch zur Entstehung der Fangblase von Utricularia analysieren. Mit freundlichen Grüßen, W.-E.L. P.S. Ich möchte bei dieser Gelegenheit auch an meine Fragen zum Thema Entstehung des Auges erinnern (Brief vom 12. 10. 99): "Zum Thema Auge möchte ich in Verbindung mit meiner Arbeit Auge widerlegt Zufalls-Evolution folgende Fragen an Sie richten:
Auszug aus meinem Brief vom 12.11.1999 und Ergänzungen:
(Vorbemerkung: Herr A. ist auf die oben genannten fünf Punkte nicht mehr zu sprechen gekommen (auch nicht auf die Fragen zum Auge). Statt dessen hatte er unter Berufung auf "eine Koryphäe auf dem Gebiet der Paläontologie und Evolutionsforschung", die nun bedauerlicherweise völlig falsch informiert war, u. a. behauptet, Brush wüsste nichts von den neuesten Saurierfunden. A. nahm also die Behauptungen der 'Koryphäe' unkritisch zur Kenntnis, um Brush als inkompetent hinzustellen. Tatsächlich aber war Brush einer der ersten Forscher, die in China selbst die neuesten Funde begutachtet haben. Daher meine Antwort:)
"Zum Thema des Ursprungs der Federn: Prof. Brush kennt natürlich die neuen Saurierfunde und hat darüber auch ausführlich referiert. Speziell in diesem Zusammenhang hat er seine Auffassung von den genetischen "Revolutionen" noch einmal bekräftigt.
Für die Familie der Federkeratingene haben wir hier am Institut eine Suche nach homologen Sequenzen in allen Datenbanken der Welt durchgeführt: Es gibt keinerlei homologe Sequenzen. D.h. nach allem was wir bis heute wissen können, sind diese Sequenzen einmalig (viele Molekularbiologen sind der Meinung, dass wir die Sequenzen von fast allen Genfamilien — etwa 4000 - der Tiere bereits kennen).
Wie also sind die Federkeratingene entstanden? Nach meinem Verständnis durch intelligente DNA-Programmierung."
Des weiteren behauptete Herr A., dass - "selbst wenn also aus dem chemischen Aufbau isoliert keine direkten Hinweise auf die Abstammung von Vögeln aus Reptilien hervorgehen sollten, lassen sich in der Summe zahlreiche Hinweise (auch zahlreiche Indizienbeweise) dafür finden" - worauf, kurz zusammengefasst, nahezu sämtliche Homologiephänomene von Herrn A. als Beweise für die generelle Evolutionstheorie aufgeführt wurden.
Ich habe darauf geantwortet mit dem Hinweis auf die Diskussion der Homologieproblematik in den einschlägigen Kapiteln des ganz ausgezeichneten Werkes von Junker und Scherer (1998): EVOLUTION - EIN KRITISCHES LEHRBUCH, sowie mit Bezugnahme auf Walter ReMine's THE BIOTIC MESSAGE (1993).
An dieser Stelle sei zunächst an das Wort des Paläontologen Oskar Kuhn erinnert:
"Die Ähnlichkeit der organischen Naturformen erklärte man durch Entwicklung, diese wieder bewies man durch die abgestufte Ähnlichkeit. Dass man hier einem Zirkelschluss zum Opfer fiel, wurde kaum bemerkt; das, was man beweisen wollte, dass nämlich Ähnlichkeit auf Entwicklung beruhe, setzte man einfach voraus und machte dann die verschiedenen Grade, die Abstufung der (typischen) Ähnlichkeit, zum Beweis für die Richtigkeit der Entwicklungsidee. Albert Fleischmann hat wiederholt auf die Unlogik des obigen Gedankengangs hingewiesen. Der gleiche Gedanke diente nach ihm wechselweise als Behauptung und als Beweisgrund.
Ähnlichkeit kann aber auch auf einen Plan zurückgehen, und....Morphologen wie Louis Agassiz, einer der größten Morphologen aller Zeiten, haben die Formenähnlichkeit der Organismen auf den Schöpfungsplan, nicht auf Abstammung zurückgeführt."
Der entscheidende Punkt ist, dass die Homologien auf allen Ebenen (von der Morphologie bis zur DNA) ganz eindeutig eine einzige gemeinsame Quelle für alle Lebensformen beweisen. Denn die homologen Ähnlichkeiten sind selbstverständlich kein Zufall! Diese einzige gemeinsame Quelle ist - nach dem Verständnis der Abstammungslehre - der gemeinsame einzellige Urahn aller Lebensformen. Nach dem Verständnis der Intelligent-Design-Movement jedoch ist die gemeinsame Quelle der eine Designer, der alle Lebensformen erschaffen hat, oder in den Worten Walter J. ReMines (1993, pp. 20, 22, 23, 37, 261, 353, 361, 383):
"Life is the product of a single designer - life was intentionally designed to resist all other interpretations of origin."
"This system of living objects was constructed by a single source (e.g., a common designer)."
"Look at the unifying message. It says life was designed as a unified body of objects. Not one single species is an outlander. All organisms are tied together by underlying similarity. A complex web of factors links all life. These similarities extend down to bio molecules. Biologic universals as DNA/RNA, proteins, amino acids, and ATP join life together at the molecular level."
"Similarity and diversity both have a role in the biotic message. Similarity makes life look like the work of a single designer, while diversity makes life difficult to explain by naturalistic purposes. Similarity and diversity are the alphabet of the biotic message."
"Message theory says life was designed as a biotic message. Life was designed to look like the product of a single designer (the unifying message). Yet life was also designed to resist evolutionary interpretation."
"Life's designer intentionally created life to look like a message from a single designer."
"Besides the nested pattern, the unity of life is augmented with various features which subtly knit life together more visibly than biochemicals can. For example, mimicry, mutualism, symbiotic relationships, and convergences unite the system of life into a unified whole. Darwin was able to see this unity even though he had no knowledge of the universalities at the biochemical level. This is one unified system of life; attributable to only one designer."
"The embryological similarities serve to directly unify life on a broader scale than can be observed by adult organisms alone. The system of life was designed to be unified, and the biomessage sender has gone to great lengths to ensure it is seen that way."
Walter ReMine hat diese Punkte in seiner BIOTIC MESSAGE ausführlich und, wie ich meine, überzeugend begründet. Der Leser prüfe bitte selbst, ob meine Aussage zutrifft. Schließlich sei noch erwähnt, dass die Botaniker und Zoologen der sogenannten "idealistischen Morphologie" diese Punkte seit spätestens J.W.Goethe im Prinzip schon genauso gesehen haben (jedoch oft nicht so explizit formulierten wie ReMine). Der Botaniker Wilhelm Troll und seine Schule seien hier besonders hervorgehoben:
"Wäre die Artbildung, wie es der Darwinismus alter und neuester Prägung will, wirklich ein richtungsloses Geschehen und würde durch sie ein Formenkontinuum geschaffen, aus dem erst sekundär, durch Ausfall der Übergangsformen, bestimmt umgrenzte Gruppen herausgeschnitten werden, so müßten gerade die Zwischenglieder über die typischen Vertreter einer natürlichen Gruppe, in unserem Beispiel der Orchideen, in der Überzahl sein, was offenbar den Tatsachen widerspricht. Nicht die stetige Reihe der Zwischenglieder ist also das typisch einigende Band und die "Einheit des Typus" bestimmt umgrenzter Formenkreise nur durch eine Störung des Formenkontinuums bewirkte täuschende Erscheinung; sondern umgekehrt: das s t e t i g e körperliche Band von Form zu Form existiert überhaupt nicht; wenn trotzdem ein inniger morphologischer Zusammenhang zwischen den Gliedern eines Formenkreises besteht, so ist das eben ein "geistiges" Band und findet seinen Ausdruck im Typusbegriff" - W. Troll 1984, p. 77 in: Gestalt und Urbild. Hrsg.: L. Behling.
Was unsere obige Diskussion zur Utricularia-Fangblase und die Vogelfeder anlangt, so wird das von ReMine explizit beschriebene Prinzip an diesen Beispielen besonders deutlich: Beide hochkomplexen Strukturen sind fest eingebunden in das Web of Life ("Life is the product of a single designer…"), und doch kann auch nach ununterbrochenen 150-jährigen Bemühungen keine überzeugende evolutionstheoretische Antwort zu ihrer Entstehung gegeben werden (" - life was intentionally designed to resist all other interpretations of origin.")
Zum Ursprung der Vögel vgl. weiter:http://www.trueorigin.org//birdevo.asp , -ein paar brauchbare Hinweise findet der Leser auch bei http://www.zillmer.com/leseprob.htm - ob die Hauptthese des letzteren Autors zutrifft - Saurier und Mensch gleichzeitig - möchte ich hier nicht diskutieren. Zu neueren Evolutionsproblemen der Vögel aufgrund der Liaoning-Funde siehe weiter: http://www.wort-und-wissen.de/sij/sij51-5.html und zu Protopteryx ("birds before dinosaurs") http://www.abcnews.go.com/sections/science/DailyNews/dinosaur_bird001207.html. Weitere Punkte: http://www.foxnews.com/scitech/112600/feathers.sml und http://www.answersingenesis.org/docs/1352.asp sowie http://creationists.org/chapter4.html. (Die Literaturhinweise - ich zitiere u.a. Materialisten und Kreationisten - implizieren nur meine Identifikation mit den zutreffenden naturwissenschaftlichen Argumenten, in der Regel jedoch nicht mit den weltanschaulichen Anliegen der Verfasser. - Zur Frage nach meiner Position zum Kreationismus vgl. Sie bitte die Ausführungen oben, die nächtsten beiden Kapitel und die Diskussion zur Stellungnahme von Herrn Prof. D..)
Die hier bis in Detail diskutierten biologischen Tatsachen sind soweit also in voller Übereinstimmung mit Mendels, von Gärtners und Linnés Schlussfolgerung, "dass der Species feste Grenzen gesteckt sind, über welche hinaus sie sich nicht zu ändern vermag."
Nachtrag 1. Juni 2001:
Einige relevante Punkte
aus der Diskussion mit Prof. J. (im Mai 2001) (Hervorhebungen im
Schriftbild von mir, außer NOT in Großbuchstaben von Prof. J.):
W.-E. L.: "I had a discussion with a very simple
minded but agressive neo-Darwinian who insisted that the bird's feathers
are absolutely nothing but frayed-out reptilian scales and that there is
hardly anything easier on this earth than to derive feathers from reptile
scales." Prof. J.: "Feathers certainly are NOT frayed-out
reptilian scales despite the strong suggestions of anatomists such as
A.S. Romer and the model of Phil Regal. Among other things it would
require a Lamarkian type of evolution! In contrast there is considerable evidence that
feathers are a novel derative of the epidermal placode. In addition
to
what we know about the feather keratins and the structural genes, there is
now evidence of major differences in ontogeny. The evolution* of
development involved several bout of expanded redundancy and introduction
of branching patterns. These mechanisms mirror the protein configuration
and other processes. Simple fraying leading to the redundancy in
branching
simply is not only inadequate, but also incorrect." W.-E.L.: "Is there anything in (a) the beta-sheet or
(b) overall construction of the phi-keratins that is principally different
from the beta-sheets of reptiles or the overall construction of reptilian
beta-keratins?" Prof. J.: "In answer to the queries in your note of
16 May: The Beta (B-) sheet of feather keratins is essentially similar in
structure (a secondary structure) to all B-sheets in proteins. They may
differ in primary sequence, but the shapes are the same. B-sheets also
occur in non-keratin proteins. The overall structure of the feather
keratin family is also quite similar. This includes feathers of all
generations and morphology, claw, beak and scale. Still, these differ
from all other B-keratins, except in their helical [beta-pleated
sheet?] conformation. The other major group of keratins have the alpha
helical conformation. All keratins have increased numbers of cysteine
residues (forming disulfide bonds), and a primary sequence that includes
many nonessential amino acids. They are quite insoluble primarily because
of the disulfide bonds and the secondary structure. Hence they must be
denatured in order to get into solution. Within the group (feather, claw-scale-beak), the
monomer unit in feathers is the smallest, about 100 amino acids with the
central B-sheets. There are no cysteines in the B-sheets, but it is rich
in prolines (at each turn) and other simple amino acids. The c-s-b protein
has essentially the same structure but it is longer and differs by a
repeating tripartite (Gly-Gly-X). Feathers have 18-24 different monomers
which differ in residues only in the globular portion. These feather
(phi) keratins are a subset of the other hard keratins...It is only a
matter of semantics how one terms these categories. Some use feather- and
non-feather-B-keratins. I find it easier and more direct to use Phi for
feathers, all else is B-. This has been discussed by Roger Sawyer
(American Zoologist vol 40, no 6)." W.-E.L.: "Many thanks for your detailed reply to my
questions. Yet, for the time being, there is one last question on the
topic in my mind: I have assumed that reptilian scales consist
mainly of
alpha-keratins. Is that true or not?" Prof. J.: "Not! reptilian scales and claws, and some
other appendages (e.g. the rattle on rattle snakes) contain a beta
keratin. But it is different from that in birds. The soft
parts of the skin of reptiles, birds and mammals contain alpha
keratins. Different genes, different filaments, etc."
Das heisst also: "In reptiles, alpha-keratins provide
[only!]
the structural proteins
of the skin and the flexible portions of scales..." (siehe
oben). Wie anfangs schon hervorgehoben, gibt es
Tausende von beta-Proteinen, die nicht voneinander ableitbar
sind, so dass die sekundäre Faltblattstruktur allein im
abstammungstheoretischen Sinne noch gar nichts beweist (die
Ähnlichkeit
dürfte
funktionale Gründe haben). Die nach Brush wiedergegebene
Aussage hingegen zur
"unterschiedlichen Proteingestalt und -sequenz" bleibt für die
Sequenz
sowieso richtig, aber auch für die Gesamtproteingestalt. Die
zahlreichen weiteren oben diskutierten Differenzen widerlegen die damit
nachgewiesenermaßen falsche neodarwinistische
"Zerschlitzungshypothese".
(24. 7. 2001:) Kommen wir kurz noch einmal zum Anfang unserer Ausführungen
zurück: Ich habe dort das Spektrum-Standardlehrbuch der BIOLOGIE 1997, p. 712, mit der Aussage
zitiert: "Federn bestehen aus beta-Keratin - demselben Protein, das auch die Schuppen der Reptilien bildet." Wir
können jetzt also feststellen: Beta-Keratin ist richtig, - nicht aber die folgende Aussage, "demselben
Protein, das auch die Schuppen der Reptilien bildet." Weiter wurden Lehninger et al. zitiert, die in ihrem PRINCIPLES OF BIOCHEMISTRY (1993,
p. 171) für die Vogelfeder alpha-Keratin angeben (das ist nun direkt falsch!), welches auch bei Reptilien
vorkommt (alpha-Keratin kommt zwar auch bei Reptilien vor, das ist jedoch für die Entstehung der Vogelfeder
irrelevant). Ein wenig verwirrend ist bei alledem noch die Nomenklatur. Wenn eine Protein weder als alpha-
noch als beta-, sondern als phi-Keratin klassifiziert wird, besteht die Tendenz, anzunehmen, dass es sich weder um
alpha noch um beta-Strukturen, sondern um eine eigene weitere Kategorie handelt ("There are three groups of
keratins: the alpha-keratins, the beta-keratins and the feather keratins", - D. Parry 1999, p. 1343
in: Encyclopaedia of Molecular Biology, Vol. 2; siehe oben). Das trifft jedoch im Falle der Proteinstruktur der
Vogelfeder nicht zu.
____
(I)Ich hatte ursprünglich geschrieben (mein im
Vorwort
und Nachtrag erwähnter und korrigierter Fehler): "Die sekundäre
Faltblattstruktur der Proteine
reptiler Schuppen
unterscheidet sich von jener der Vogelfedern auf frappierende Weise
(phi-Keratine der Federn im Unterschied zu den
(meist) alpha-Keratinen der Reptilienschuppen). Gleiches trifft auf die Elementaranalyse zu." - Eine genaue
Elementaranalyse dürfte jedenfalls bei stark unterschiedlichen Proteinen auch zu unterschiedlichen
Ergebnissen
führen. *Im Gegensatz zu seiner
naturwissenschaftlich gut begründeten Widerlegung der
"Zerschlitzungshypothese", sind Prof. J.s eigene Evolutionshypothesen
auch nicht mehr als Denkmöglichkeiten bzw. logische
Postulate
unter der Voraussetzung der Evolutionstheorie oder einfach aufgrund
gegebener Strukturen (so musste z.B. beim Erstellen des
genetischen Programms der
Federbildung unter anderem auch die Information für "introduction of
branching
patterns" generiert worden sein) -
der rein mechanistische Ansatz durch die bekannten Zufallsmutationen
aber
ist
unbewiesen (wie er selbst in unserer Diskussion einräumte).