Herr M. B. schrieb mir aus Costa Rica am 4. Mai 2021 u.a. zu weiteren inzwischen erfolgten Untersuchungen und Forschungserbebnissen:
"....bei mir wachsen gerade 8 Stck. 'flower spikes' an 2 großen Coryanthes-Pflanzen.
Ref. (1) http://www.weloennig.de/CorCat.html
Ref. (2) Thomas Eltz (2005) 'Juggling with Volatiles'
Ref. (1) erwähnt 'Hummeln die ölige Substanzen aufnehmen' als Bestäuber von Coryanthes-Blüten ... Das ist falsch, denn die Bestäuber sind Bienen der Gattung Euglossa und diese nehmen nicht ölige Substanzen' auf, sondern Düfte, siehe ref. (2).
Es ist offensichtlich nicht hilfreich daß Sie deutsche Fachleute aus alter Vergangenheit zitieren, um Ihre Einwände zu Darwin's Evolutionstheorie deutlich zu machen (smile) ..."
Aus meiner Antwort vom 7. Mai 2021
Hallo Herr M. B.:
Vielen Dank für Ihre Mail. Freut mich sehr, von Ihnen zu hören, dass Sie sich als Hobby-Gärtner mit dieser wirklich 'fantastischen' Orchidee Coryanthes beschäftigen ("bei mir wachsen gerade 8 Stck. 'flower spikes' an 2 großen Coryanthes-Pflanzen").
Sie haben recht: Charles Darwin hat sich mit seinen Hinweisen zu den "Hummeln" in Verbindung mit Coryanthes total geirrt ("eine Anzahl großer Hummeln" ... "eine 'ununterbrochene Prozession' von Hummeln"... "das die Hummeln am Fortfliegen verhindert"). Das war 1877.
Aber wieso sprechen Sie in diesem Zusammenhang von "deutschen Fachleuten" (smile)?
Aber Wolfgang Kuhn (1984) hätte es besser wissen müssen (auch wenn er kein Orchideen-Fachmann war).
Richtig bleibt jedoch sein wissenschaftlicher Einwand gegen den Darwinismus.
Die zitierten Autoren Meeuse und Morris liegen dagegen (1984) mit der folgenden Aussage völlig richtig ("... selten anzutreffenden Prachtbienenmännchen - manchmal sogar zu Dutzenden. Diese Bienen, die die Coryanthes-Blüten besuchen, gehören zu den schönsten Insekten, die es gibt; sie weisen - je nach Art - glänzende, irisierende Grün-, Blau- oder Bronzetönungen auf."
Siehe auch die systematischen Details in https://en.wikipedia.org/wiki/Euglossa)
Bastiaan Meeuse war Professor für Botanik an der University of Washington.
Sean Morris: Oxford Scientific Filmmakers
Diese Fachleute Meeuse und Morris formulieren jedoch [1984 noch] ungenau, wenn sie schreiben (von mir jetzt der Kern-Text kursiv):
"Zunächst landen die Bienen noch nicht auf den Blüten, sondern fliegen nur höchst aufgeregt umher,
wobei sie sich gelegentlich in der Luft gegenseitig rammen, als wollten sie miteinander um den Besitz der
von der Orchidee angebotenen Substanzen wetteifern. Nach ein paar Minuten beginnen sie, am Rand,
an den Außenwänden und auf dem pilzförmigen Gebilde (es sitzt auf der den zwei Drüsen gegenüberliegenden Seite
des Randes) zu landen. Alle kratzen mit ihren bürstenähnlichen Vorderfüßen ein paar Sekunden
lang wie wild an der wachsartigen Oberfläche der Blüte. Dann ziehen sie sich ein Stückchen zurück,
schweben in der Luft und verstauen die gesammelten Ölsubstanzen in Taschen an ihren Hinterbeinen.
Danach landen sie wieder auf der Blüte und wiederholen den ganzen Vorgang." Wo genau sammeln die Bienen
"the waxy substance containing the scent" (Formulierung in der Quelle siehe unten).
"The floral fragrance is produced by osmophores hidden under the mostly cup-shaped hypochile of the lip.
These volatile substances are both the attractant and reward for the bees" (siehe wieder unten). "The flower secretes a fluid (see Coryanthes alborosea picture) into the flower lip,
which is shaped like a bucket. The male orchid bees (not the females)
are attracted to the flower by a strong scent from aromatic oils,
which they store in specialized spongy pouches inside their swollen hind legs,
as they appear to use the scent in their courtship dances in order to attract females.
The bees, trying to get the waxy substance containing the scent,
sometimes fall to the fluid-filled bucket. As they are trying to escape, they find that there
are some small knobs on which they can climb on, while the rest of the lip is lined with smooth, downward-pointing hairs,
upon which their claws cannot find a grip. The knobs lead to a spout (see the Coryanthes leucocorys picture), but as the bee is trying to escape, the spout constricts. At that same moment, the small packets containing the pollen of the orchid get pressed against the thorax of the bee. However, the glue on the pollen packets does not set immediately, so the orchid keeps the bee trapped until the glue has set.
...The bee, having stored the aromatic oils in his back legs, can then fly off to mate with a female bee."
https://en.wikipedia.org/wiki/Coryanthes
Aus der Antwort von Herrn M. B. am 7. Mai 2021: Vorbemerkung: Die folgende Abhandlung stammt zum größten Teil
aus einer
Diskussion zwischen Herrn Dr. A. und mir vom 1. 12. 1995; der Text wurde
erweitert, insbesondere durch ein längeres Zitat zu Catasetum nach Meeuse und Morris und Darwins
Abbildungen zu beiden Orchideenarten sowie Fußnoten etc. 2002 (Hervorhebungen im
Schriftbild - außer
Artnamen - der Zitate fast alle von mir). - Einige Punkte würde ich
heute wohl etwas "weniger
grimmig" formulieren, aber die Tatsachen und die Argumentation bleiben von dieser stilistischen Frage
unberührt. Zusätzlich sei noch angemerkt, dass ich Darwins Motivation zu seinem Orchideenwerk
ausgesprochen aufschlussreich finde. An Asa Gray schrieb er: "[N]o one else has perceived that my chief interest
in my orchid book has been that it was a 'flank movement' on the enemy" (zitiert nach H. Cronin 1993, p. 26, bold von mir).
"The enemy" waren natürlich die 'defenders of deliberate design'.
Der um Wahrheit ringende Leser urteile bitte beim gründlichen Studium der folgenden Diskussion selbst, ob Darwins Angriff erfolgreich war oder nicht: Zur Ergänzung zu Utricularia möchte
ich noch zwei weitere Beispiele gegen den Darwinismus und die Synthetische Evolutionstheorie aufführen. Es
handelt sich um die erstaunlichen Blütenstrukturen der Orchideengattungen Coryanthes und
Catasetum, die Charles Darwin vor mehr als 125 Jahren in seiner ORIGIN wie folgt beschrieben hat
(zunächst zu
Coryanthes - die Abbildung vorweg stammt aus Darwins Orchideenabhandlung 1877; Darstellungen nach
neueren Befunden siehe unten, ebenso Darwins Catasetum-Abbildungen): Diese Orchidee
hat ihr Labellum (Unterlippe) teilweise zu einem großen Gefäß ausgehöhlt, in das
fortwährend aus zwei darüber befindlichen Hörnern Tropfen fast reinen Wassers herabsickern. Ist das
Gefäß halb voll, so fließt das Wasser durch einen
seitlichen Abguß ab. Der Basalteil der
Unterlippe steht über dem Gefäß und ist selbst kammerartig ausgehöhlt mit zwei
Seiteneingängen; in dieser Kammer befinden sich sonderbare fleischige Leisten. Der intelligenteste Mensch
könnte, wenn er es nicht sähe, unmöglich ausfindig machen, welchem Zwecke all diese Teile
dienen. Crüger sah jedoch, daß eine Anzahl großer Hummeln
die Riesenblüten der Orchideen
besuchten, nicht um Nektar zu schlürfen, sondern um die Leisten in den Kammern oberhalb des
Gefäßes abzunagen. Während sie dies taten, stießen sie einander häufig in das
Gefäß; dadurch wurden ihre Flügel so naß, daß
sie nicht fortfliegen konnten und durch den vom
Ausguß gebildeten Durchgang kriechen mußten. Crüger sah
eine
"ununterbrochene Prozession" von Hummeln in
dieser Weise aus dem unfreiwilligen Bade kriechen. Der Durchgang ist eng und hat ein von Säulen getragenes
Dach, so daß die Hummel beim Durchzwängen ihren Rücken
erst an der klebrigen Narbe reibt und dann an
den klebrigen Drüsen der Pollenmassen. Diese kleben dadurch am Rücken derjenigen Hummel fest, die
zufällig zuerst durch den Gang einer eben entfalteten Blüte kriecht, und werden so
fortgetragen. Crüger sandte mir in Spiritus eine Blüte mit einer Hummel, die er getötet hatte,
bevor sie ganz herausgekrochen war; auf ihrem Rücken befand sich noch die Pollenmasse. Fliegt nun eine so
belastete Hummel zu einer andern Blüte oder zum zweitenmal zu derselben zurück, wird sie von ihren
Genossen in das Gefäß gestoßen, und kriecht sie dann durch den Gang, so kommt die Pollenmasse mit
der klebrigen Narbe in Verbindung, bleibt hier hängen und - die Blüte ist befruchtet. Jetzt erst
erkennen wir den vollen Wert jedes einzelnen Blütenteils: der Wasser absondernden Hörner und des
halbgefüllten Gefäßes, das die Hummeln am Fortfliegen verhindert und sie zwingt, durch die
Öffnung zu kriechen und sich an den zweckmäßig aufgestellten klebrigen Pollenmassen und der
klebrigen Narbe zu reiben. Der Bau der Blüte einer verwandten Orchidee
(Catasetum)# ist vollkommen anders, obgleich er demselben Zweck dient, er ist aber ebenso
merkwürdig. Wie die der Coryanthes, so werden auch ihre Blüten von Bienen besucht, die das
Labellum benagen. Dabei berühren sie unweigerlich einen langen, spitzzulaufenden empfindlichen Fortsatz, den
ich Antenne genannt habe. Diese Antenne überträgt bei der Berührung eine Empfindung oder Schwingung
auf eine gewisse Membran, die sofort aufbricht, und dadurch wird eine Feder ausgelöst, die die Pollenmasse
wie einen Pfeil vorschnellt und ihr klebriges Ende an den Rücken der Biene heftet. Die Pollenmasse der
männlichen Pflanzen (diese Orchideen sind getrenntgeschlechtlich) wird demnach auf die Blüte der
weiblichen Pflanze übertragen, wo sie mit der Narbe in Berührung kommt, die genügend klebrig ist,
um gewisse elastische Fäden zu zerreißen und die Pollenmasse zurückzubehalten; damit ist die
Befruchtung bewirkt. Nach diesen
Ausführungen, sollte man ja nun erwarten, dass uns der Autor zu seinem Thema The Origin of Species
eine überzeugende Erklärung dafür liefert, wie sich aus 'normalen' Orchideen Schritt für
Schritt eine Coryanthes- bzw. eine Catasetum-Orchidee
entwickelt haben soll: Wie viele und
welche Schritte waren dafür notwendig? Welche Selektionsvorteile hatten die einzelnen Schritte
(Veränderungen) gegenüber dem Bau der herkömmlichen Orchideen? Durch welche genetischen
Ursachen [wie wir heute sagen würden - man kann auch generell nach den Ursachen fragen] wurden die
angenommenen vielen kleinen Schritte bewirkt? Wie sieht
der experimentelle Beweis für die Richtigkeit dieser Annahmen aus? Keine einzige dieser Fragen
wird jedoch von Darwin gestellt und/oder beantwortet. Statt dessen folgen einige (ziemlich
nichtssagende) generalisierende Feststellungen über Variabilität und Vererbung 'modifizierter Bildungen'
und dass die Natur keine Sprünge macht (natura non facit saltum) und dass sehr viele kleine
Schritte für die Bildung neuer Organe notwendig waren ("denn die natürliche Zuchtwahl wirkt nur
dadurch, daß sie sich kleine allmähliche Abänderungen
zunutze macht; sie kann nie einen großen
und plötzlichen Sprung machen, sondern muss mit kurzen und sicheren, aber langsamen Schritten
voranschreiten" - Darwin 1876, p. 224). Er räumt immerhin ein, dass es "in vielen Fällen sehr schwer
auch nur zu mutmaßen ist, durch welche Übergänge viele Organe zu ihrer jetzigen Beschaffenheit
gelangt seien", doch behauptet er direkt anschließend, dass selten ein Organ vorkommt, von welchem man
keine hinleitenden Übergangsstufen kennt usw. Nun könnte man natürlich
nicht ganz unberechtigt darauf hinweisen, dass mit Darwin die Evolutionstheorie noch ziemlich am Anfang war (wenn
auch Lamarck und St. Hilaire schon 70 Jahre zuvor evolutionäre Ideen und Theorien veröffentlicht
hatten) und er die Antworten auf die entscheidenden Fragen damals noch nicht wissen konnte. Inzwischen sind jedoch
mehr als 100 Jahre intensivster Forschung auf allen Gebieten der Biologie vergangen (mit den beiden oben
erwähnten Autoren fast 200 Jahre). Wo stehen wir also heute mit diesen Fragen? Kann nun die moderne
Synthetische (oder irgendeine andere) Evolutionstheorie die Fragen zu den oben zitierten Beispielen
überzeugend beantworten? Meines Wissens trifft das nicht zu! Statt dessen gewinnen
grundsätzliche Einwände gegen den darwinschen Ansatz in den
letzten Jahren immer mehr an Bedeutung: Der bedeutende Botaniker Karl Goebel bemerkte zu den
Orchideenblüten, "daß alle die
wunderbaren Anpassungen der Orchideenblüten nicht mehr erreichen, als zahllose viel einfachere
Bestäubungseinrichtungen. Wir finden die verwickelten Einrichtungen vieler Orchideenblüten,
z.B. Catasetum mit seiner Pollinienabschleuderung, Coryanthes mit ihrem Lippenbad für die
Blütenbesucher, als Kunstwerk wundervoll. Aber vom platten Nützlichkeitsstandpunkt aus ist das alles
eine "Luxusanpassung", die uns dann verständlich ist, wenn sie für die Formen, welche sie aufweisen,
eine durch ihre ganze Organisation bestimmte und gegebene war. Dann ist
sie [die 'Anpassung'] kein Luxus, sondern eine zwar nicht an
sich (d.h. wenn man nur auf das Rücksicht nimmt, was erreicht wird), aber für eine
Orchidee zweckmäßige. Die Selektionstheorie weist eine solche durch innere Ursachen in
bestimmter Richtung erfolgende Gestaltung ab. Aber sie ist weder zureichend, die "Entstehung der Arten" noch die
Vielfältigkeit der Anpassungen uns verständlich zu machen: eben weil sie der Morphologie den
"logos" nimmt und sie in ein Gemenge an sich richtungslose[r] Variationen
auflösen will" (bold/kursiv von Goebel gesperrt). Wilhelm Troll, der größte Pflanzenmorphologe des 20. Jahrhunderts, referiert und
kommentiert Goebel u.a. wie folgt: "... vom platten Nützlichkeitsstandpunkt aus seien Einrichtungen wie die
einer Catasetum-Blüte mit ihrer Pollinienabschleuderung oder einer Coryanthes-Blüte mit
ihrem Lippenbad für die Blütenbesucher Luxusanpassungen, die nicht mehr erreichen als die viel
einfacheren Bestäubungseinrichtungen zahlreicher anderer Orchideen und deshalb im Kampf ums Dasein
nicht gezüchtet sein können..." Professor Wolfgang Kuhn
(Universität Saarbrücken) bemerkt zu Coryanthes: Immerhin räumt der alte Darwin im
Anschluß an dieses Paradebeispiel gegen
den blinden Zufall als "schöpferische Ursache" ein, es sei "in vielen Fällen äußerst
schwierig...auch nur zu mutmaßen, durch welche Übergänge Organe ihren jetzigen Zustand erreicht
haben". Nur denkt er leider, wie seine Nachfolger noch immer, nicht konsequent bis zu Ende. So stellt sich ihm
erst gar nicht die Frage, wie denn die hypothetischen "Vorstufen" eines derartigen "Präzisionsmechanismus",
einer solch fehlerfrei funktionierenden "Ganzheit", wie man heute sagen würde, im angeblich erbarmungslosen
Kampf ums Dasein hätten überleben können. Immerhin waren sie ja während dieser Entwicklung
noch nicht "fertig", mithin also unvollkommen und damit nicht "funktionsfähig". Die "natürliche
Auslese" hätte demnach leichtes Spiel damit gehabt, sie auszumerzen! Es wäre für einen
Wahrscheinlichkeitsmathematiker sicher eine reizvolle, wenn auch trotz aller modernen Computer, die er mit den
entsprechenden Daten füttern könnte, kaum lösbare Aufgabe, die Wahrscheinlichkeit oder, treffender
gesagt, die zweifellos nicht einmal mehr in astronomischen Zahlen auszudrückende Unwahrscheinlichkeit eines
"zufälligen" Zusammentreffens aller zum richtigen Funktionieren nun einmal erforderlichen Voraussetzungen
auszurechnen - und zwar "auf beiden Seiten": bei der Blüte sowohl als auch bei ihren Besuchern, den
Hummeln! Max Thürkauf hat diese enorme Unwahrscheinlichkeit einer rein zufälligen
Entstehung von "Synorganisation", jener planvollen Zuordnung, die jeden lebendigen Organismus kennzeichnet, einmal
treffend karikiert. Es sei, so bemerkte er, sehr viel wahrscheinlicher,
daß eine Katze, die auf den Tasten einer
Schreibmaschine herumhüpft, dabei zufällig Goethes Faust (erster
und zweiter Teil!) schreibt, als daß
ein so komplizierter Organismus wie der einer Katze jemals durch Zufall zustandekäme! Um Ihnen noch einmal ein Bild von der
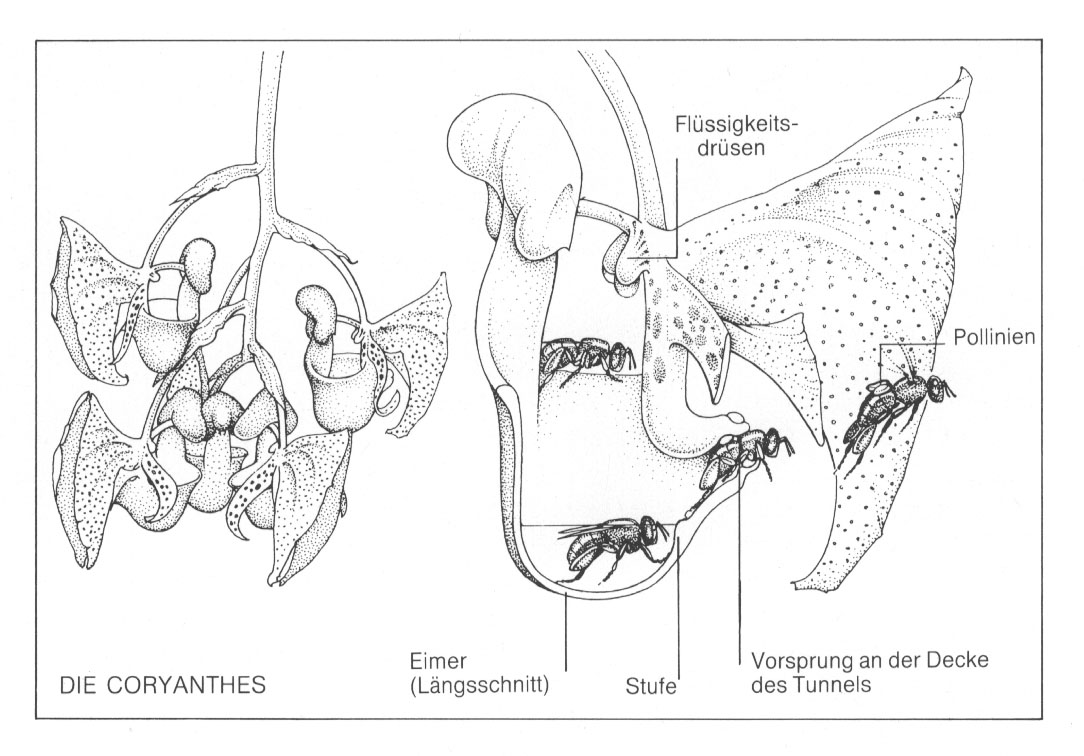
Komplexität der Coryanthes-Blüte zu vermitteln, habe ich Ihnen im folgenden einige Abbildungen
und weitere Zitate zu diesem Thema zusammengestellt: Blütenstand von Coryanthes speciosa, Helmorchidee. Abbildung aus Peter Arnold (1994, p. 32; Orchideen):
Die folgende Zeichnung von Coryanthes-Blüten und des Längsschnitts durch eine Blüte
samt Bewegungsablauf einer Prachtbiene aus B. Meeuse und S. Morris 1984, p. 59: Meeuse und
Morris bemerken zu dieser Darstellung: "Männliche Prachtbienen sammeln in den Blüten dieser
Coryanthes-Art ein öliges Sekret. Dabei verlieren sie oft den Halt und purzeln in den "Eimer". Die von
der Flüssigkeit durchnäßte Biene findet nur einen einzigen
Fluchtweg - einen günstig gelegenen
Tunnel an der Seite des Kessels. Am Ende des Tunnels jedoch hält die Pflanze sie fest, klebt ihr zwei
Pollenpakete auf den Rücken und hält sie so lange gefangen, bis der Klebstoff fest geworden ist. Sobald
die Biene sich befreit hat, kann sie wieder eine andere Coryanthes-Orchidee besuchen, wo sich im Grunde
genommen der ganze Vorgang wiederholt; doch diesmal nimmt ihr ein Haken am Ende des Tunnels geschickt die
Pollenpakete wieder vom Rücken." Weiter
beschreiben Meeuse und Morris mehr als 100 Jahre nach Darwins Schriften zum Ursprung der Arten und über die
Orchideen die
Blütenstrukturen von Coryanthes sehr anschaulich wie folgt (pp. 58-60): Der Aufbau der Blüte ist - selbst für den
Botaniker, der
Überraschungen gewohnt ist - äußerst erstaunlich. Die Sepalen, die sich wie gefaltete
Schmetterlingsflügel zurückbiegen, enthüllen ein Gebilde, das einem steilwandigen, unbedeckten
Eimer ähnelt. Am Rand des Eimers wächst ein pilzförmiges Objekt nach oben. Über der Eimermitte
hängen zwei rundliche Verdickungen - Drüsen, die in den ersten paar Stunden nach Öffnung der
Blüte ständig eine klare Flüssigkeit absondern. Diese tropft hinunter in den Eimer. Nach ein paar
Stunden ist die Blüte ungefähr 6 Millimeter hoch mit Flüssigkeit gefüllt. Dann werden die
Drüsen "abgestellt". In die Seite des Eimers mündet - gerade über dem Flüssigkeitsspiegel -
ein kurzer Tunnel, der dem Schnabel einer Gießkanne ähnelt. Am Eingang dieses Tunnels befindet sich
eine teilweise von der Flüssigkeit bedeckte, kleine "Beule" in der Eimerwand. An der Decke des
Trichterausgangs sitzen Pollinien und Narbe der Blüte. Die Blüten sind - je nach
Art - gelb oder gelb mit braunen und orangefarbenen Flecken. Der Beobachter, der herausfinden will, wie dieses
kunstvolle Blütengebilde "funktioniert", braucht normalerweise gar nicht lange zu warten. Schon während
der Minuten, in denen sie sich öffnet, strömt die Blüte einen schweren, seifenartigen,
süßen Duft aus. Wie durch ein Zauberwort herbeigerufen, erscheinen die normalerweise etwas scheuen,
selten anzutreffenden Prachtbienenmännchen - manchmal sogar zu
Dutzenden. Diese Bienen, die die Coryanthes-Blüten
besuchen, gehören zu den schönsten Insekten, die es
gibt; sie weisen - je nach Art - glänzende, irisierende Grün-,
Blau- oder Bronzetönungen auf. Zunächst
landen die Bienen noch nicht auf den Blüten, sondern fliegen nur
höchst aufgeregt umher, wobei sie sich gelegentlich in der Luft
gegenseitig rammen, als wollten sie miteinander um den Besitz der von der
Orchidee angebotenen Substanzen wetteifern. Nach ein paar Minuten beginnen
sie, am Rand, an den Außenwänden und auf dem pilzförmigen
Gebilde (es sitzt auf der den zwei Drüsen gegenüberliegenden
Seite des Randes) zu landen. Alle kratzen mit ihren
bürstenähnlichen Vorderfüßen ein paar Sekunden lang
wie wild an der wachsartigen Oberfläche der Blüte. Dann ziehen
sie sich ein Stückchen zurück, schweben in der Luft und
verstauen die gesammelten Ölsubstanzen in Taschen an ihren
Hinterbeinen. Danach landen sie wieder auf der Blüte und wiederholen
den ganzen Vorgang. Wenn bis zu zwölf Bienen
an jeder Blüte eifrig Orchideensubstanzen sammeln und sich dabei
gleichzeitig auch noch gelegentlich in Luftgefechte miteinander einlassen,
ist es kein Wunder, daß früher oder später eine den Halt
verliert und in die Mitte der Blüte fällt. Sie geht in der
klaren Flüssigkeit sofort unter. (Experimente haben gezeigt,
daß die Flüssigkeit ein Benetzungsmittel enthält und daher
eine nur sehr geringe Oberflächenspannung aufweist.) Die Biene
bemüht sich freizukommen, doch die Wände sind wachsähnlich
und glatt und so kommt sie nicht weiter. Sie würde in der
Flüssigkeit rasch ertrinken, wenn es nicht einen in diese
Blütenfalle eingebauten Fluchtweg gäbe - den
Tunnel. Während die Biene verzweifelt herumzappelt, kommen ihre
Vorderfüße mit der "Beule" an der Seite des Eimers in
Berührung. Diese Beule ist eine Art Treppenstufe, welche die
durchnäßte Biene in den Tunneleingang führt. Der Tunnel
schließt sich eng um die Biene, und sie muß sich
buchstäblich zum Ausgang vorkämpfen. Doch gerade in dem
Augenblick, in dem es schon so aussieht, als werde sie nun endlich ihre
Freiheit wiedererlangen, drückt sich ein an der Tunneldecke
befindlicher Vorsprung fest in den Spalt zwischen ihrem Brustkasten und
ihrem Hinterleib. Obwohl sie sich energisch wehrt, wird sie festgehalten,
und die Blüte klebt ihr ihre Pollinien fest auf den
Rücken! Wodurch die Dauer der Gefangenschaft einer solchen Biene
bestimmt wird, wie der Mechanismus in Gang gesetzt und wie die Biene dann
anschließend wieder freikommt, weiß man nicht. Nach
ungefähr zehn Minuten kommt die mit den Pollinien beladene Biene
schließlich zum Vorschein und fliegt davon, um zu trocknen. Was geschieht in der Zwischenzeit mit den anderen Bienen,
die fleißig ihre Orchideensubstanz sammeln? Viele von ihnen erleiden
das gleiche Schicksal wie die erste Biene - nur mit dem einen Unterschied,
daß sie nun, da die Pollinien schon entfernt sind, ungehindert durch
den Fluchttunnel kriechen können wie Besucher durch ein Drehkreuz. Es
werden weiterhin neue Bienen angelockt, bis die Blüte bestäubt
ist oder sie am Ende ihres zweiten Tages verwelkt - je nachdem, was eher
eintritt. Zur Bestäubung kommt es, wenn eine Biene, die Pollinien von
einer anderen Coryanthes-Pflanze am Leib trägt, in die
Blüte stürzt. Diese Biene muß sich auf ihrer Flucht an
einem an der Tunneldecke befindlichen Greifhaken vorbeidrängen; und
dieser pflückt ihr die Pollinien vom Rücken. Die Chance, daß das geschieht, ist jedoch zwangsläufig sehr gering. Die Coryanthes ist keine verbreitete Orchidee und blüht außerdem in unregelmäßigen Abständen. Die Bienen sind auch nicht gerade häufig, obwohl eine blühende Coryanthes angeblich über eine Entfernung von bis zu 8 Kilometern hinweg Bienenmännchen anzieht, werden - wenn überhaupt - nur wenige von ihnen gerade Pollinien von einer anderen Pflanze auf dem Rücken tragen. Außerdem fällt ja auch nicht jeder Besucher in den Eimer. Die Chancen für diese Orchidee scheinen also ausgesprochen schlecht zu stehen. Beobachtungen der Coryanthes in freier Natur ergaben, daß die Pflanzen nur sehr selten mehr als eine Samenkapsel pro Blütenstil trugen - in den meisten Fällen gar keine. Wenn es jedoch zu einem solchen "Glückstreffer" kommt und Bestäubung erfolgt, so enthält die entstehende Samenkapsel Hunderttausende mikroskopisch kleiner Samen, die mit dem Wind davontreiben, wenn die reife Samenkapsel schließlich aufspringt und sie freiläßt.
Abbildung aus Meeuse und Morris, p. 60.
Als Kommentar zur Abbildung bemerken Meeuse und Morris: "Ein Pollinienpaar ist
fest an den
Hinterleib dieser Prachtbiene gekitted - diese Traglast hat sie sich bei einer früheren Begegnung mit einer
Coryanthes-Orchidee erworben. Während ihrer Flucht aus der mit Flüssigkeit gefüllten Falle
dieser zweiten Blüte werden der Biene die Pollenpakete vom Rücken gepflückt. So kommt die
Fremdbestäubung dieser Orchideen zustande." - Siehe weiter http://www.pbs.org/wnet/nature/orchids/deceivers.html
(mit Movie; Voraussetzung mindestens QuickTime 3). Ein Serie ganz hervorragender Aufnahmen von 34 Coryanthes-Spezies sowie Aufschluss über
weitere funktionale Strukturen wie den extrafloralen Nektarien der Gattung findet
der daran interessierte Leser bei Günter Gerlach, Konservator am Botanischen Garten München,
THE SPECIES OF THE GENUS CORYANTHES.
G. Gerlach beschäftigt sich seit zwanzig Jahren mit der Gattung.
Sehen wir uns jetzt Catasetum einmal etwas genauer an. Die folgenden beiden Aufnahmen
wurden uns freundlicherweise von Herrn Dr. G. Gerlach am 22. 10. 2002 zur Verfügung gestellt: Auf der Abbildung links sind männliche Blüten und auf der Abbildung rechts eine weibliche Blüte
mit einem Prachtbienenmännchen dargestellt.
Zu Catasetum
schreiben Meeuse und Morris pp. 61/62 ("Unterthema Prachtbienen und Catasetum-Scharfschützen"): Die vielen südamerikanischen Orchideenarten der
Gattung Catasetum bedienen sich bei der Bestäubung eines "Scharfschützen"-Mechanismus. Hier ist
die Sache jedoch noch komplizierter, denn nur die männlichen Blüten wenden diese Technik
an: Catasetum ist eine der wenigen Orchideen, bei denen man getrennte männliche und weibliche
Blüten findet - für gewöhnlich an verschiedenen Pflanzen. Sie unterscheiden sich im Aufbau so sehr,
daß man kaum glauben möchte, daß sie zu ein und derselben Art gehören. Beide sehen
allerdings ziemlich unscheinbar aus, erzeugen jedoch einen starken Duft, der bei den männlichen Exemplaren
zwei oder drei Tage nach der Öffnung auftritt, bei den weiblichen dagegen erst mehrere Tage später. Die Bestäuber sind
Prachtbienenmännchen... [die ebenfalls zur Gattung Euglossa
gehören; Anmerkung von W.-E.L.] Sie sind ganz begierig danach, den Duftstoff zu sammeln, der von dem
Drüsengewebe
in der Lippenhöhlung der Blüte erzeugt wird. Wenn die Tiere in die Duftkammern der männlichen
Blüten eindringen, können sie fast gar nicht umhin, eine oder beide der scharfen Spitzen eines Paares
höchst merkwürdiger Anhängsel zu berühren. Diese werden als "Römer" oder
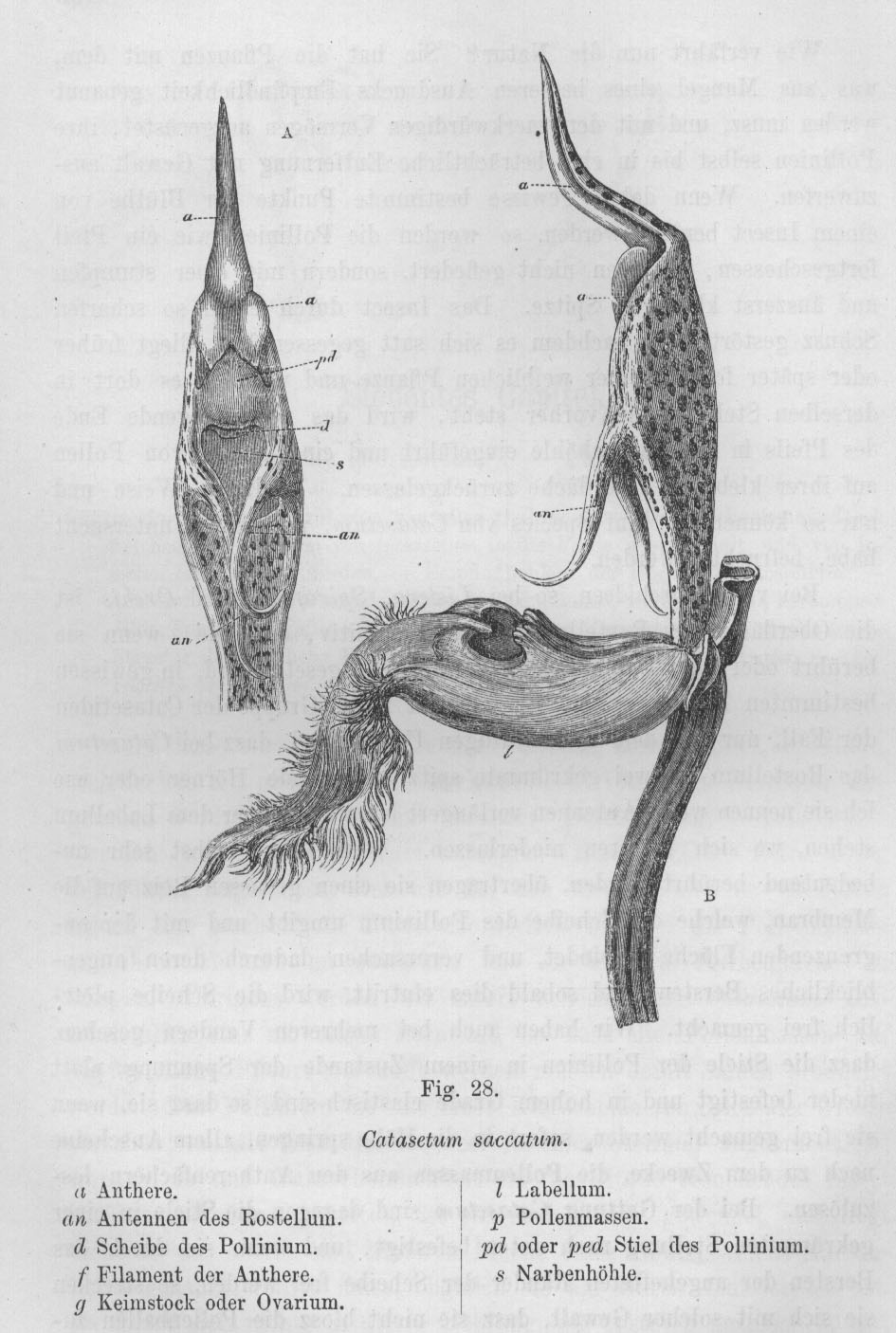
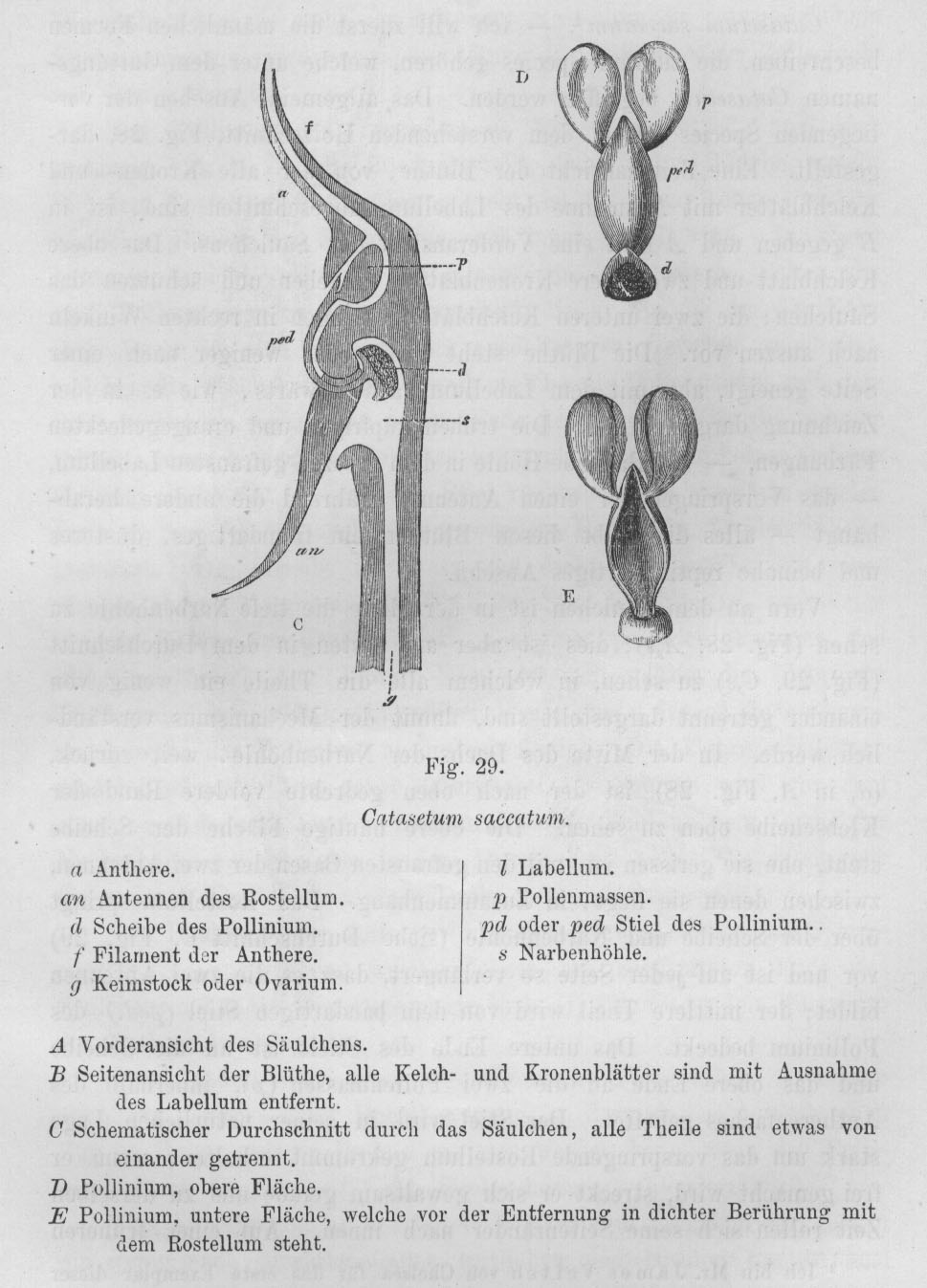
"Antennen" bezeichnet und ragen aus jenem Teil der Blüte hervor, in dem die Pollinien verborgen sind. Durch
einen dünnen, elastischen Stiel sind die Pollinien mit der sogenannten Klebscheibe verbunden. Diese befindet
sich in einem kleinen Sack, und zwar so, daß der Stiel unter Spannung in gekrümmter Position gehalten
wird. Schon bei einer leichten, durch die zarte Berührung einer Biene hervorgerufenen Biegung der Hörner
wird die Klebscheibe mit großer Kraft aus ihrem Sack befreit, denn der gebogene Stiel der Pollinien springt
hoch wie eine Metallfeder. Das ganze Pollinarium (die beiden Pollinien mit ihrem Stiel und der Klebscheibe) wird
herausgeschleudert, macht einen halben Salto rückwärts und trifft - mit der Klebscheibe zuerst - die
Biene. Der Aufprall ist so stark, daß das Insekt manchmal sogar aus der Blüte herausgeschlagen
wird! Experimente haben gezeigt, daß das Pollinarium - wenn die Biene dieses Geschoß nicht mit ihrem
Körper abfinge - 80 Zentimeter weit fliegen würde. Da ist es kaum verwunderlich, daß die
Klebscheibe sofort fest an dem Insekt haften bleibt! Die Treffsicherheit dieser Blüten ist geradezu
unheimlich; nur welcher spezielle Körperteil der Biene jeweils getroffen wird, das ist von Art zu Art
unterschiedlich. Sobald das Pollinarium abgeschossen ist, beginnt der Geruch der Blüte
sich zu verändern, und nach kurzer Zeit ist er ganz weg. Die männliche Blüte hat nun ihre Aufgabe
erfüllt. Die weiblichen Blüten beginnen ihren Duft erst einige Tage später zu erzeugen, und sie
strömen ihn auch nicht ununterbrochen aus, sondern an mehreren Tagen hintereinander jeweils nur ein paar
Stunden lang (für gewöhnlich ein bis zwei Stunden am Morgen). Sie duften mindestens solange, wie sie
unbestäubt sind, und das kann bis zu einem Monat dauern. Bei einem
pollinariumbeladenen Prachtbienenmännchen, das in die Duft-Höhle einer weiblichen Blüte kriecht,
baumelt das Pollinarium unter dem Körper. Wenn das Insekt die Blüte dann, mit Duftstoffen beladen,
wieder verläßt, bewegt es sich dabei rückwärts; und wenn diese Catasetum zur selben
Art gehört, von der auch das Pollinarium stammt - aber nur dann -, sind die Pollinien genau in der
richtigen Position und bleiben in der das befruchtungsbereite Narbengewebe bildenden Rinne hängen. Hier
bleiben sie dann, während Stiel und Klebscheibe abbrechen und von der Biene davongetragen werden. Die
Blüte ist bestäubt. Binnen einer halben Stunde verliert sie ihren Duft, und von jetzt an kümmert
sich keine einzige Biene mehr um sie. Jede Catasetum-Art hat ihren eigenen
besonderen Geruch und zieht daher auch nur ihre eigenen besonderen Euglossa-Bienen
an. Außerdem funktioniert der Schleudermechanismus mit derartiger Präzision, daß die
Pollinien immer nur an einen bestimmten Körperteil des Bestäubers angeheftet werden: nämlich den
für die Bestäubung "richtigen". Infolgedessen kommt Hybridisation zwischen verschiedenen
Catasetum-Arten in freier Natur nur äußerst selten vor. Catasetum-Abbildungen aus Darwin 1877, pp. 154/155.
Prof. Klaus Lunau - ein Befürworter der Synthetischen Evolutionstheorie
- vermerkt zu C. fimbriatum und weiteren Arten unter anderem (2002, p. 89): Während des Besuchs der staminaten Blüte löst das Prachtbienenmännchen
beim Parfümsammeln einen Triggermechanismus aus, der das Pollinium wie mit einem Katapult an den Körper des
Prachtbienenmännchens schießt, wo das Pollinium mit einer Klebscheibe haftet. Die Geschwindigkeit des abgeschossenen
Polliniums wurde bei Catasetum fimbriatum auf 323 cm/sec bestimmt. Die derart beschossenen Prachtbienenmännchen
verlassen darauf fluchtartig die Blüte - mit Pollinien. Beobachtungen von Gustavo Romero und Craig Nelson (1986) an
Catasetum ochraceum ergaben, dass von 86 an C. ochraceum beobachteten Prachtbienenmännchen kein einziges
zum zweiten Mal eine männliche Blüte dieser Art aufsuchte. Offensichtlich reicht in diesem Fall eine einzige
derartige Erfahrung aus, dass sie diese Signalkonstellation nicht mehr anfliegen. Auf diese Weise hat diese staminate Catasetumblüte
alle männliche Konkurrenz aus dem Feld geschlagen und gleichzeitig die Wahrscheinlichkeit für einen Folgebesuch einer
ähnlich duftenden, aber völlig anders aussehenden pistillaten Blüte erhöht.
Nach K. Lunaus Hypothese imitieren die von den Parfümorchideen erzeugten Duftöle
'Duftstoffe der für die Weibchen attraktiven Harze, die diese zum Nestbau verwenden'. Außerdem imitierten die weiblichen
Catasetum-Blüten einen Prachtbienennesteingang. - Der Verfasser macht jedoch keinen Versuch, die Entstehung der Synorganisationen bei
Catasetum im Sinne seiner Theorie Schritt für Schritt zu erklären (siehe auch die weiteren Anmerkungen unten). Kommen wir [damit] auf die Frage nach der Entstehung solcher
Systeme zurück: Wie oben ausgeführt,
hatte weder Darwin eine überzeugende evolutionäre Erklärung für die Entstehung solcher
synorganisierten Strukturen, noch haben die zeitgenössischen Evolutionisten eine naturalistische
Erklärung. Evolutionstheoretiker können zur Zeit nur glauben,
dass alles "von
selbst" entstanden ist, aber sie können es nicht beweisen. In
gänzlicher
Abwesenheit einer überzeugenden wissenschaftlichen Beweisführung zur selektionstheoretischen Entstehung
der
genannten Blütenstrukturen der Orchideen versucht Darwin seine Theorie zu retten, indem er just in diesem
Zusammenhang die Schöpfungstheorie [seiner biologischen und weiteren Zeitgenossen] angreift. Die dahinter
stehende Taktik ist: Versuche, die Schöpfungstheorie als
unglaubwürdig hinzustellen und der Leser wird
die Evolutionslehre akzeptieren. [Wären die folgenden Argumente Darwins gegen Intelligent Design
tatsächlich zutreffend, dann hätte diese Methode als einzige Wahl zwischen zwei Möglichkeiten - und
das wird dabei vorausgesetzt - sogar eine gewisse Berechtigung; aber sehen wir uns seine Einwände einmal
etwas genauer an]. Wir lesen: Es ist
gewiß richtig, daß neue Organe sehr selten oder nie
plötzlich bei einem Wesen* erscheinen, als ob sie
für einen besonderen Zweck geschaffen worden wären; - wie es auch schon die alte, obwohl etwas
übertriebene naturgeschichtliche Regel "Natura non facit saltum" anerkannt wird. Wir finden dies in den
Schriften fast aller erfahrenen Naturforscher angenommen: MILNE EDWARDS hat es treffend mit den Worten
ausgedrückt: Die Natur ist verschwenderisch in Abänderungen, aber geizig in Neuerungen. Warum sollte es
nach der Schöpfungstheorie so viel Abänderung und so wenig wirklich Neues geben? (Darwin
p. 223/224). Wenn das richtig
wäre, dann würde das ja mit anderen Worten für die Schöpfungstheorie bedeuten, dass der
Schöpfer "verschwenderisch in Abänderungen, aber geizig in Neuerungen" wäre. Und ein in
unwesentlichen Dingen verschwenderischer und obendrein in allen wesentlichen Neuerungen geiziger
Schöpfer ist ja wohl für keinen vernünftigen Menschen und Wissenschaftler akzeptabel und
zumutbar. Also lassen wir die Schöpfungstheorie fallen und akzeptieren wir die Evolutionslehre, die ja trotz
ihrer Erklärungsschwierigkeiten zumindest nicht solche unvernünftigen Annahmen zu machen braucht. Genauer betrachtet irrt Darwin in allen wesentlichen Punkten: Es ist schlicht und
einfach falsch, "daß neue Organe sehr selten oder nie
plötzlich bei einem Wesen erscheinen, als
ob sie für einen besonderen Zweck geschaffen wären". Bei praktisch allen neuen Tierstämmen, die
beispielsweise im Kambrium auftreten, erscheinen die neuen Organismen der Tierstämme mit sämtlichen
Organen so plötzlich und abrupt, dass selbst Darwin zugab (p. 396): "Diese Tatsache muss vorerst
unerklärt bleiben und wird mit Recht als eine wesentliche Einrede gegen die hier entwickelten Ansichten
hervorgehoben werden." Und diese Tatsache ist bis heute nicht nur unerklärt, sondern durch viele weitere
Fossilfunde für die Evolutionslehre noch viel schwieriger geworden. Ein [umfangreiches Werk] könnte man
zum plötzlichen Erscheinen neuer Organe verfassen. (Das schließt nicht aus, dass Organe nach ihrem
plötzlichen Auftreten bei den ersten Organismen später bei der Verwirklichung weiterer und zum Teil auch
komplexerer Formen noch weiter ausgestaltet werden - vergleichen Sie bitte meine Schrift über das Auge, wo
ich in allen Einzelheiten in Verbindung mit diesem Beispiel auf diese Fragen zu sprechen gekommen bin. Jedenfalls
ist auch solche Ausgestaltung häufig mit völlig neuen und plötzlichen Schritten verbunden,
etwa die Linsenbildung, das Farbensehen, die Bildung der Tränendrüsen etc.) Die Behauptung "Natura non facit saltum" ist in unserem Jahrhundert von der Quantentheorie
bis zur Mutationsforschung so gründlich widerlegt worden, dass wir den Satz umkehren können: Die Natur
macht nur Sprünge. "Wir finden
dies [den Kontinuitätsgedanken] in den Schriften fast aller erfahrenen Naturforscher angenommen:" Die größten Naturforscher wie
Linné und Cuvier (beides Schöpfungstheoretiker) und ihre Schulen waren da offensichtlich anderer
Meinung. Es handelt sich dabei vielmehr um eine philosophische Grundhaltung als um eine naturwissenschaftliche
Erkenntnis. "Milne Edwards hat es treffend
mit den Worten ausgedrückt: Die Natur ist verschwenderisch in Abänderungen, aber geizig in
Neuerungen." Die Fülle
der verschiedenen Baupläne (der großen Neuerungen) im Tier- und Pflanzenreich ist so groß, dass
ich keinen einzigen Biologen nennen kann, der alle diese Baupläne kennt, versteht und in der Lage wäre,
sie alle aus dem Stehgreif zu erklären! Aber auch an kleinen Neuerungen
(innerhalb der Baupläne) besteht kein Mangel, und Abänderungen gibt es natürlich auch in
Fülle. Dass die Zahl der Abänderungen die der Neuerungen übertrifft liegt in der Natur der
Sache. Ein Beispiel aus der Technik: Autos sind fast alle nach dem gleichen Prinzip gebaut: Ottomotor meist vorne
(selten hinten wie beim alten VW-Käfer), in der Regel besitzen sie vier Räder, Bremsen, Sitzbänke,
Scheiben ringsherum, und ohne Lenkrad geht es nicht etc. etc.. Völlige Neuerungen sind
recht selten, aber die Abänderungen sind fast uferlos. Jedes Modell hat seine Abänderungen und wenn man
genauer hinsieht, meist auch einige zumindest kleinere Neuerungen. Ähnlich verhält es sich mit
Hubschraubern und anderen Fluggeräten, U-Booten, Segelschiffen, Computern usw.: Die Möglichkeiten der
Abänderungen sind fast unendlich - die technisch-funktionellen Möglichkeiten völliger Neuerungen
aber erscheinen eher begrenzt (die Möglichkeiten der kleineren Neuerungen - d.h. innerhalb der
"Baupläne" - sind jedoch auch sehr groß). Welcher
vernünftige Mensch würde jedoch im
Prinzip eine
Argumentation wie die folgende akzeptieren: "Der Mensch ist verschwenderisch in Abänderungen, aber geizig in
Neuerungen. Warum sollte es nach der Autoschöpfungstheorie so viel Abänderung und so wenig wirklich
Neues geben?" - Also sind Autos nicht gebaut ("erschaffen") worden, sondern durch Evolution von selbst
entstanden! Wir wissen zwar nicht wie, wo und warum sich die Automobile von selbst gebildet haben, auch fehlt der
experimentelle Beweis, aber wir wollen glauben, dass das so ist.** Was ich hiermit
zeigen möchte, ist die Tatsache, dass sämtliche evolutionstheoretischen Erklärungsversuche von
Darwin bis heute an den Realitäten vorbeigehen. Catasetum und Coryanthes und tausend andere
Phänomene sind bis heute nicht evolutionstheoretisch erklärt. Nach meinem Verständnis sind sie nur durch Intelligent Design [in
Verbindung mit der Message Theory] erklärbar als Ausdruck der
Weisheit und Macht und Genialität ihres
Designers - und manchmal, so möchte ich hinzufügen, wohl auch
für einen gewissen Sinn
für Humor. [Zu grundlegenden Einwänden zur Schlussfolgerung auf Intelligent Design
vgl. Artbegriff und zu Fragen der
Systematik vgl. weiter
das Thema Evolution und Klassifikation.] Jede
simple Konstruktion in der Technik setzt einen Konstrukteur voraus, wie
viel mehr
die tausendmal komplexeren Konstruktionen in der Tier- und Pflanzenwelt! Ich weiß natürlich, dass diese
Schlussfolgerung für jeden Materialisten blanke Ketzerei ist. Dafür ist sie jedoch wahr. Selbstverständlich darf man anderer Meinung sein. Wir haben hier in der Bundesrepublik
Deutschland glücklicherweise Religions- und Glaubensfreiheit (grundgesetzlich garantiert). Man kann die
Evolutionslehre zu einem unantastbaren religiösen Dogma erheben und grundsätzlich keinerlei
naturwissenschaftliche Argumente gegen sie gelten lassen. Unter solchen Voraussetzungen wäre es dann aber
auch nicht sinnvoll, über dieses Dogma naturwissenschaftlich zu diskutieren. Jede naturwissenschaftliche
Theorie hingegen muss in der Lage sein, Falsifikationskriterien zu nennen,
sonst ist sie keine wissenschaftliche
Theorie. Für die Intelligent-Design-Theorie habe ich solche in der Diskussion mit Prof. K. genannt. Nun hätte ich natürlich auch von Ihnen sehr gern Falsifikationskriterien für Ihre
Evolutionstheorie gehört. Grundsätzlich will ich mich auch von der Richtigkeit
der Evolutionstheorie überzeugen lassen. Ich lade Sie daher ein, mir für Utricularia, Catasetum
und Coryanthes die folgenden (auf pp. 3 und 4 schon genannten und jetzt etwas
übersichtlicher aufgeführten) Fragen zu beantworten: 1) Wie viele und welche
morphologisch-anatomischen und physiologischen Schritte waren für die Bildung der jeweils neuen Strukturen
und Anordnungen zur
Entwicklung von Coryanthes und Catasetum erforderlich? 2) Durch welche
genetischen Ursachen wurden die angenommenen vielen kleinen (oder wenigen großen?) Schritte
bewirkt? Können die uns bekannten Zufallsmutationen tatsächlich eine wissenschaftlich ausreichende
Erklärung für die Entstehung dieser synorganisierten Strukturen bieten? (Vgl. die Details zur
Mutationsfrage: Mutationen: Das
Gesetz der rekurrenten Variation sowie
Genduplikationen und Artbegriff)***. 3) Welche Selektionsvorteile hatten die einzelnen Schritte sowohl auf morphologisch-anatomischer
als auch auf genetischer Ebene im Vergleich zu den jeweiligen Ausgangsformen bzw. den ursprünglichen
Wildtypgenen ("Vorstufen")? 4) Wie sehen die experimentellen Beweise für die
Richtigkeit der neodarwinistischen Behauptungen aus? [In seinem Werk Evolutionary Biology von 1998 zitiert Douglas J. Futuyma
beide Orchideenarten und gibt auch Darwins Abbildungen von 1877 dazu wieder, aber Futuyma macht nicht einmal den Versuch einer
evolutionstheoretischen Erklärung! Statt dessen bemüht er sich darum - wie schon einst Darwin -, Schöpfung bzw.
Intelligent Design als prinzipiell unmöglich hinzustellen. - Der oben zitierte Klaus Lunau hingegen erwähnt keinerlei Alternativtheorien
zum Neodarwinismus. Bei der Frage nach der schrittweisen Entstehung der Synorganisationen bei den Orchideen setzt er die Richtigkeit der
Synthetischen Evolutionstheorie einfach voraus. Der Verfasser bleibt aber sehr allgemein (um es vorsichtig zu formulieren), wenn er nach
Beschreibung der Pollinien u.a. erklärt (2002, p. 88): "Die starke Reduktion der notwendigen Anzahl von Blütenbesuchen, bei dem
im Extremfall der gesamte Einsatz (Pollen) auf einen Blütenbesuch gesetzt wird, ist für die Orchideen offenbar ein
Spiel mit gutem Ausgang." Da die Orchideen in großer Formenfülle existieren (1 040 Gattungen und je nach Schätzung 15 000 bis 20 000 Spezies),
war das Mutations-/Rekombinations- und Selektionsspiel mit den zitierten Reduktionsphänomenen "offenbar ein Spiel mit gutem Ausgang"
- denn sonst gäbe es sie ja nicht! Weiter schreibt der Verfasser: "Orchideen konnten dadurch Bestäubungsstrategien entwickeln,
die im Pflanzenreich einmalig sind." - Auch hier wird die Synthetische Evolutionstheorie wieder undiskutiert als richtig voraussetzt,
aber der aufmerksam-kritische Leser sieht sehr schnell,
dass damit noch keine einzige der oben aufgeführten und für die Theorie entscheidenden Fragen beantwortet ist
(und weiter: wie konnten sich Pflanzen mit einer starken Reduktion der Anzahl von Blütenbesuchen etc. gegenüber anderen ohne
Reduktion behaupten,
bevor sie die dazu gehörigen, einmaligen Bestäubungsstrategien 'entwickelt' hatten?).
Da in der umfassenden und in der biologischen Tatsachenbeschreibung sowie grafischen Gestaltung
gelungenen Arbeit von K. Lunau (2002) zur Mimikrythematik und verwandten Themen die Synthetische Evolutionstheorie die ausschließliche
Grundlage zur Beantwortung der Frage nach der Entstehung der Strukturen bildet und da Letzteres ebenso auf viele weitere neuere und ältere Arbeiten
zutrifft (für eine recht umfassende ältere Arbeit siehe Wickler 1968), wollen wir uns zunächst einmal zur grundsätzlichen Problematik der Selektionstheorie noch einige weitere Details
genauer ansehen.
Oben hatte ich schon mit Karl Goebel und Wilhelm Troll erwähnt, dass "Einrichtungen wie die einer
Catasetum-Blüte mit ihrer Pollinienabschleuderung oder einer Coryanthes-Blüte mit ihrem Lippenbad für die Blütenbesucher
Luxusanpassungen [sind], die nicht mehr erreichen als die viel einfacheren Bestäubungseinrichtungen zahlreicher anderer Orchideen und deshalb im Kampf
ums Dasein nicht gezüchtet sein können".
David Attenborough bemerkt nach Hinweis auf die in Madagaskar heimische Orchidee
Angraecum sesquipedale sowie den sie bestäubenden Nachtfalter in ähnlicher Weise (1995, pp. 104/106) : Auch andere Orchideen sind dafür bekannt, sehr komplizierte und aufwändige
Bestäubungssysteme entwickelt zu haben, wobei man sich in einigen Fällen fragen muß warum soviel Energie investiert wurde,
um ein Resultat zu erzielen, das die meisten Pflanzen mit sehr viel weniger Aufwand erreichen.
Das gilt etwa für die Helmblume [Coryanthes], die in den Wipfeln der Urwaldbäume Mittelamerikas wächst.
Der oben schon zitierte Wolfgang Kuhn hat sich mit der Selektionsfrage zu den
Orchideen wie folgt weiter auseinander gesetzt (1985, Bd. 2, pp. 177-182), wenn er zwar in populärwissenschaftlicher Sprache, aber
mit höchst bedeutsamem wissenschaftlichem Inhalt Folgendes ausführt (Hervorhebungen, außer kursiv "müssen", im Schriftbild wieder von mir):
Überraschenderweise jedoch scheint es gerade niemanden zu überraschen,
daß es unleugbar mehr Gänseblümchen und Löwenzahnpflanzen, ja noch weit mehr viel einfacher gebaute "Butterblumen" auf
unseren Wiesen gibt als Orchideen. Sie alle vermochten demnach im "Kampf ums Dasein" zu bestehen und zu überleben - sogar besser und sehr,
sehr viel erfolgreicher als die paar Orchideen, die hier und da versteckt mitten unter ihnen blühen mögen. Da ihre Nachkommenzahl
also außerordentlich viel größer ist als bei diesen ausgesprochenen, schutzbedürftigen Seltenheiten, erwiesen sie sich
nach Darwin als die bei weitem "Tüchtigeren", die "Fittesten" im "Kampf ums Überleben". Weshalb also hätten sich jemals Orchideen
entwickeln müssen, wo doch gerade dieser evolutive Prozeß ihre Überlebenschancen stark verringerte, statt sie im Gegenteil zu erhöhen?
Man könnte es freilich auch einfacher formulieren: Warum denn in aller Welt so kompliziert, wenn doch just das gleiche rascher, erfolgreicher
und so unendlich viel einfacher funktioniert?
Die Synthetische Evolutionstheorie behauptet jedoch nicht, dass die Orchideen mit Notwendigkeit
entstehen "mussten", sondern völlig ungeplant in dem oben genannten "Spiel" eben auch entstanden seien.
Man fragt sich jedoch, wie die Orchideen im von Kuhn genannten Widerspruch zu diesen Spielregeln der Selektion überhaupt entstehen konnten
(- von den niemals beobachteten, vergleichbare Strukturen aufbauenden Mutationen einmal
ganz abgesehen [ich möchte nur einmal eine aus Mutationsexperimenten mit D. melanogaster hervorgegangene,
wesentliche größere Fliege mit klarer schwarzgelber Warnstreifung mit Überlebenschancen in der Natur sehen] - es sei an dieser
Stelle noch einmal an den Beitrag
Mutationen:
Das Gesetz der rekurrenten Variation erinnert.) - Kuhn fährt fort:
Eine ganz gewöhnliche "Butterblume" - welche unserer vielen Hahnenfußarten
der Volksmund auch immer so bezeichnen mag - ist nicht nur am Wegrand oder Wiesenrain, auf der Waldwiese ebenso wie auf dürftigem Sandboden,
auf trockenen Kalkhängen oder sogar im feuchten, nährstoffarmen Hochmoorboden überall "zuhause", während die Insekten-Orchideen
ausschließlich auf Kalkböden mit ganz bestimmten PH-Werten angewiesen sind. Sie brauchen darüberhinaus auch weder raffinierte
Düfte zu erzeugen, noch irgendwelche Insekten nachahmende Attrappenformen auszubilden, um dennoch bestäubt zu werden.
Ihnen genügt einfaches gelbes Pigment durchaus, um den Insekten vor dem grünen Hintergrund der anderen Wiesenpflanzen aufzufallen,
und sie können, um ihnen ausreichend Pollen mitzugeben, auf "Pollinien" mit "Regenschutz" und Haftmechanismen absolut verzichten!
Nicht einmal irgendwelche Saftmale als "Wegweiser" für die Blütenbesucher sind erforderlich. Winzige Mengen Nektar
am Grunde der gelben Nektarblätter (die wie Blütenkronblätter aussehen) unter bergenden Schuppen abgesondert,
belohnen die bestäubenden Insekten für ihre Mühe. Aber sogar das ist völlig überflüssiger "Luxus",
denn bei einigen Hahnenfußarten fehlen selbst diese Nektardrüsen - und auch sie sterben keineswegs etwa aus
im "Kampf ums Dasein"!
Im Grunde genommen hätte es die Evolution
(von der seltsamerweise manche Neodarwinisten wie von einer Person reden!) also ruhig bei der einfachsten und zugleich erfolgreichsten Blütenform
belassen können, etwa bei sternförmigen, "radiärsymmetrischen" Blüten, wie sie heute noch die Hahnenfußgewächse,
besonders auffällig etwa das häufige Scharbockskraut oder das im Frühjahr gleichzeitig mit ihm erscheinende Buschwindröschen
besitzt. Es wäre absolut nicht "nötig" gewesen, andere Blütenformen auszubilden, wie sie beispielsweise
die Lippenblütler, Rachenblütler, Glockenblumen usw. besitzen. Wiederum die Zahl der Nachkommen als Maßstab angelegt,
haben sie sich keineswegs durch diese unvermeintlichen "Verbesserungen" im Blütenbau als die "Tüchtigsten" erwiesen!
Reinhard Eichelbeck schreibt zur Selektionsfrage des für unsere Orchideen relevanten
Themas der Mimese und Mimikry (1999, pp. 195/196):
Und eine weitere Frage drängt sich auf: Wenn Mimese oder Mimikry tatsächlich
einen derartigen Überlebensvorteil haben, warum gibt es sie dann nicht öfter? Diese Fragestellung nenne man in Amerika den
"Kühlschrankirrtum", meinte Professor Wolfgang Wickler, Verfasser eines Standardwerkes über Mimikry,
in einem Artikel der Zeitschrift "Natur": "Denn natürlich kann dasselbe Argument auf Kühlschränke angewendet werden.
Kühlschränke können keinen so großen Vorteil bieten, wie man gemeinhin annimmt, denn die meisten Menschen auf der Welt kommen
ohne Kühlschrank aus." Der Logik dieses Argumentes folgend, könnte man sagen: Der Kühlschrank ist in der Tat zwar bequem,
aber kein unbedingter "Überlebensvorteil", denn der Mensch hat es schließlich in der Vergangenheit geschafft, mehr als 50 000 Jahre
ohne Kühlschränke zu überleben. Und wenn für Mimese und Mimikry das gleiche gilt wie für den Kühlschrank,
dann sind sie also auch nicht unbedingte "Überlebensvorteile", sondern sie sind eben einfach nur bequem. Wie kann das aber sein,
wo doch nach Darwin alles, was von der "natürlichen Selektion" erzeugt wird, einen "Überlebensvorteil" im
"Kampf ums Dasein" darstellen muß?
Diese Erklärung wäre jedenfalls denkbar weit entfernt
von der Selektionsbegründung Darwins und der heutigen Synthetischen Evolutionstheorie, ganz abgesehen davon, dass
die neuen Synorganisationen äußerst "unbequem" zu sein scheinen.
Darüber hinaus haben Wicklers Populationen mit den Kühlschränken
im allgemeinen eine geringere Nachkommenschaft als die ohne und wären damit auf lange Sicht zum Aussterben verurteilt!
Diese Anmerkungen bedeuten jedoch nicht, dass bei der Thematik der
Mimese und Mimikry die
Selektionsfrage insgesamt irrelevant sei. Es gibt Beispiele für frequency-dependant selection verschiedener
Erscheinungsbilder in Populationen einer Art und die
oft nur noch als
"phantastisch" zu bezeichnenenden Tarn- und Warntrachten im Organismenreich haben für die betroffenen Spezies in aller Regel
auch Überlebensfunktionen (man vergleiche nur einmal
die eindrucksvollen Beispiele, die u.a. in dem großformatigen Buch von Ferrari (1997)
oder auch bei Lunau (2002) und Wickler (1968) dargestellt sind.
Der entscheidende Fehler vieler Autoren liegt jedoch darin, dass sie mit diesen Funktionen auch den Modus ihrer Entstehung durch Mutation und Selektion
ohne weitere Diskussion als gegeben voraussetzen. Das "Survival of the Fittest" erklärt jedoch noch nicht das "Arrival of the Fittest",
wie kürzlich ein bestens informierter Autor mit einer sehr starken Präferenz für neodarwinistische Erklärungen eingeräumt hat.
Wenn selbst ein scheinbar so vollkommen klar und einfach liegender Fall für
mutations- und selektionsbedingte Tarnung wie der des Birkenspanners in den letzten Jahren in größte Schwierigkeiten geraten ist
(vgl. Judith Hooper [2002]: Of Moths and Men: Intrigue, Tragedy & the Peppered Moth; Fourth Estate, 377 pp.,
sowie Industriemelanismus), so darf man sich wohl fragen,
wie es dann bei genauerer Betrachtung um die große Mehrheit der weitaus komplexeren Fälle bestellt sein wird
(Mutation und Selektion können in bestimmten Phasen der Existenz von Populationen ein Rolle spielen,
aber diese Faktoren reichen nicht aus, um den Ursprung der zahlreichen Phänomene komplexer Mimese und Mimikry
in seinen Ursachen völlig zu erklären).
Kommen wir zu unseren Orchideen zurück, an denen sich diese Einwände deutlich
veranschaulichen lassen. Robert Nachtwey bemerkt in seinem ganz
ausgezeichneten Werk INSTINKT RÄTSEL DER WELT zum Aufbau und Funktion der Orchisblüten (1950, pp. 133/144):
Diese Blumen sind...der Ausdruck eines in sich vollendeten technischen Planes.
Jede Einzelheit erscheint als notwendiges Glied des Ganzen. Würde sich der Nektar frei im Sporn ansammeln,
wie es doch bei unzähligen gespornten Blüten, bei Hahnenfußgewächsen und Rachenblütlern üblich ist,
so wäre nicht die unbedingte Gewähr dafür gegeben, daß das Insekt sein Köpfchen fest in die Blüte preßt;
deshalb bleibt der Nektar im Gewebe der Spornwandung aufbewahrt. Die Narbe liegt aber dicht unterhalb des Schnäbelchens,
so daß die Pollinien notwendig auf die Narbenfläche treffen müssen, sobald sie sich nur etwas nach vorn senken.
Und daß sie sich wirklich senken, daß sie tatsächlich diese Neigung ausführen, ergibt sich wieder aus
anderen Einzelheiten der Konstruktion. Die Pollinienstielchen bestehen nämlich aus einer wasserhaltigen Gallertmasse,
die unter gewöhnlichen atmosphärischen Bedingungen schnell ihre Feuchtigkeit und ihre Spannung verliert, so daß
die Stielchen am Insektenkopf schon in einer halben Minute nach unten sinken. Am Ort ihrer Entstehung müssen sie also feucht aufbewahrt werden,
sie ruhen deshalb wohlverpackt in Säckchen, die sie vorzüglich gegen das Vertrocknen schützen. Natürlich
müssen diese Schutzbeutelchen wiederum ganz außerordentlich zart sein, damit sie sofort der Länge nach aufreißen,
wenn das Insekt die Pollinien mit sich zieht.
Die Klebscheibchen an den Pollinienstielchen tragen einen flüssigen Leim,
der an der Luft augenblicklich erhärtet und dann außerordentlich fest am Insektenkopf haftet. Auch die beiden Pollenklümpchen
sind wieder äußerst zweckmäßig eingerichtet. Sie bestehen aus vielen einzelnen Blütenstaubpäckchen,
die untereinander durch zarte, klebrige Fäden verbunden sind. Werden sie nun auf die Narbenfläche einer Blüte gedrückt,
so bleiben nur einige Pollenpäckchen auf der Narbe haften, da die Verbindungsfädchen zerreißen.
Die andern Päckchen aber bleiben am Insektenkopf, werden weiter mitgenommen und auf andere Blüten getragen.
Auf diese Weise kommen stets ungemein viele Pollenkörner aus den verschiedensten Blüten auf eine einzige Narbe,
und somit ist vielfache, wechselseitige Fremdbestäubung gewährleistet. Die sehr zahlreichen Samen eines Fruchtknotens entwickeln
dann ganz verschiedene Individuen.
Wieviel sinnvolle Erfindung in einer einzigen Orchisblüte,
jede kleinste Einzelheit hat ihre besondere Bedeutung im Plan des Ganzen und keine dürfte fehlen.
Würde der Leim am Klebescheibchen nicht sofort hart werden, so wäre alles andere sinnlos, weil die Pollinien verloren gingen.
Neigte sich das Pollinienstielchen nicht alsbald nach unten, so kämen die Blütenstaubkörner nicht auf die Narbe der neuen Blüte,
und wieder wäre der ganze Apparat vollständig umsonst gebaut.
Wenn aber jemand meint, man könnte die Entstehung all dieser
sinnvollen Einzelheiten der Orchisblüte mit der Zufallstheorie Darwins erklären,
so muß die Kraft seines Glaubens Berge versetzen können, denn ein solches zufälliges Zusammentreffen wäre
das größte Wunder.
Die Errichtung der Orchisblüte ist im Daseinskampf ebensowenig notwendig wie die Kesselfalle des Aronstabs,
denn Tausende anderer Pflanzen, die solche Apparate nicht besitzen, erhalten ihre Art und vermehren sich massenhaft.
D. h. also: Wenn auch im Vergleich zu anderen Pflanzenformen eine selektionistische
Deutung der Entstehung der neuen Funktionspläne der Orchideen nicht überzeugend gegeben werden kann, so sind doch die
geschlossenen Funktionskreise der Bauelemente für das Überleben dieser Formen sehr wohl notwendig. Die Entstehung der Gesamtsysteme ist jedoch ebenso wenig
durch Mutation und Selektion zu erklären wie die Entstehung der Kühlschränke durch ziellose Faktoren in W. Wicklers Veranschaulichung oben.
Das Survival of the Fittest betrifft in selektionsstringenten Umwelten nur die Möglichkeiten und Grenzen der Variabilität innerhalb der
Funktionskreise, wobei gleichwertige (neutrale) Variabilität bei der Bildung morphologischer Arten in vielen Tier- und Pflanzengattungen eine
erhebliche Rolle spielen dürfte. Die ganz spezifische Zuordnung von Orchideen- und Insektenarten
("The pollinators of the different Coryanthes species fit exactly to the size of the tunnel formed by the lip and the column......
only the appropriate pollinator is attracted." vgl. die weiteren Details bei
G. Gerlach) ist
damit aber noch nicht erfasst. Unter weniger selektionsstringenten Umweltverhältnissen kann es bei vielen Lebensformen aber auch zu
Degenerationserscheinungen kommen, soweit zumindest nicht grundlegend-lebensnotwendige Funktionen betroffen sind
(vgl. das Kapitel
DEGENERATION IM ORGANISMENREICH der Artbegriffsarbeit), bzw. keine alternativen Überlebensstrategien zur Verfügung stehen
(etwa eine ausreichende vegetative Fortpflanzung). Ob (und falls ja, inwieweit) die Degenerationsfrage auch auf bestimmte funktionale Strukturen
von Orchideen zutreffen könnte, ist ein noch weitgehend offenes Problem. Bei der die Orchideen kennzeichnenden Höchstspezialisierung
ist jedenfalls zu erwarten, dass bei Ausfall essentieller Funktionskreise das ganze System zusammenbricht, d.h. keine Nachkommen mehr erzeugt
werden. - Siehe darüber hinaus auch die Ausführungen der Fußnote** zum Thema "Vervollkommnung".]
Wenn jedoch die heutigen Evolutionstheorien die grundsätzlichen Fragen zum Ursprung neuer
synorganisierter Strukturen und Systeme nicht beantworten
können und trotzdem mit großer Selbstsicherheit wissenschaftliche Akzeptanz fordern, dann ist das
für mich eine reine Glaubensforderung, die auf demselben Niveau steht wie etwa die Glaubensforderung
der russisch-orthodoxen oder katholischen etc. Kirchen an deren jeweilige Dogmen. [Schlussbemerkungen: Die hier begonnene Diskussion zu den Orchideen könnte man auf
zahlreiche weitere synorganisierte Strukturen, Einrichtungen und Funktionen dieser Pflanzenfamilie ausdehnen. -
Herr Dr. A. hat keinen
der ihm seinerzeit übersandten Punkte (d.h. etwa 80% der vorliegenden Argumentation) aufgegriffen und im
Sinne einer Evolutionstheorie
beantwortet.
Zur Alternativfrage kommentiert Robert Nachtwey 1950, p. 144:
Die Orchisblüte kann nicht aus der
zufälligen Summierung vieler zusammengewürfelter Einzelheiten,
sondern nur aus der planmäßigen Zusammenfügung ihrer Teile entstanden sein.
Nach wie vor gilt in vielen Punkten auch für Coryanthes und Catasetum
Prof. Adolf Portmanns Resümee zum Ursprung der von ihm diskutierten Orchideen Gorytes und Ophrys
(1970, in der Reihenfolge pp. 535, 545, 547 und p. 542, Anmerkungen in Klammern von mir):
Der Weg ihrer "Evolution" ist in keinem Fall durch Etappen unserem Verständnis
[im Sinne des Reduktionismus] zugänglich.
Die Zweifel an der allgemeinen Geltung dieses Weges [der funktionalen Morphologie]
zum Verstehen sind aber nicht leicht zu nehmen. Der Nachweis von Strukturen, die über das funktionsgemäße Ziel hinausschießen,
die man deshalb auch als luxurierend, als hypertelisch bezeichnet hat, muß uns jeder allzu weitgehenden
funktionalen Deutung gegenüber zurückhaltend stimmen.
So, wie die Leistungssteigerung durch die Kybernetik nicht etwa die Erfindung des Neuen
im menschlichen Dasein erklärt, so erklären auch die bedeutsamen Entdeckungen der Genetik nicht das Auftreten der komplexen Neuerungen,
die über das hinausgehen, was die uns bekannten Prozesse genetischer Veränderung uns bisher vor Augen stellen.
Die Entdeckung einer für alle Lebewesen einheitlichen Struktur,
einer zur Selbstreplikation fähigen Überträgerin von Information von Schriftcharakter, hat wesentliche Konsequenzen.
Und auf der Seite 543 schreibt Portmann: "Die besondere Struktur dieser besonderen
Informationsprozesse, die uns den Vergleich mit unserer schriftlichen Verständigung aufdrängen,
lenken den Blick auf die Phänomene, die wir im menschlichen Bereich als geistige Beziehung kennen,
die ja auch bei uns nicht auf das wache Bewußtsein beschränkt ist.
Die Diskussion um das Problem des Geistigen erhält neue Impulse."
Gemäß der oben aufgeführten Argumentation gibt es zahlreiche
wissenschaftliche Gründe, die folgenden Worte des Paläontologen Oskar Kuhn und des Botanikers Wilhelm Troll zum
Typusbegriff auch auf den Subtypus der Orchideen und die Genera Coryanthes und Catasetum anzuwenden: "Der Typus ist wie jede Gestalt, realmorphologisch gesehen,
Verwirklichung eines Planes in indifferenter Materie, die ebensogut eine andere Gestalt hätte annehmen können. Als ein Universale ist der
Typus objektiv gültig, in ihm wird das Wesen der Dinge gefaßt. Naturwissenschaftlich ist die Entstehung der Typen das größte
Rätsel wegen ihres inadaptiven Charakters. Daher meinen wir mit W. Troll, daß Typen als »die Gedanken einer aus dem Welthintergrunde
in die Natur hereinwirkenden, schöpferischen Macht« aufzufassen sind, welche »der Materie neuartige Ausprägungen verleihend, die
typenhaften Grundformen ins Dasein rief«."
Vgl. weiter
Die Synthetische Evolutionstheorie und die Intelligent-Design-Theorie: Ein Vergleich (Diskussion des Haupteinwandes zu Coryanthes und Catasetum).]
____________
[1]"Gesichert" im Sinne der heutigen Möglichkeiten und Grenzen der Naturwissenschaft.
#Kurz ein Hinweis zur heutigen Systematik dieser Orchideen:
Familie: Orchidaceae
Tribus: Maxillarieae
Subtribus: Stanhopeinae
Gattung: Coryanthes (mit ca. 15 Arten, nach Gerlach inzwischen 34 Arten)
Tribus: Cymbidieae
Subtribus: Catasetinae
Gattung: Catasetum (mit ca. 50 Arten)
Die Maxillarieae sind morphologisch-systematisch mit dem Cymbidieae so nahe verwandt, dass die
Letzteren in der
Regel auf die Maxillarieae in systematischen Werken folgen. - Zur Ableitung der Namen: "Coryanthes, f.,
Gatt. der Orchidáceae (griech. kórys=Helm; ánthos=Blume; nameltlich Bezug auf die
helmförmigen unpaaren hinteren Perigonblätter des inneren Kreises; danach: Helmorchidee
(-stendel);..." (Schubert/Wagner 2000, p.167). [Der Begriff Perigon wird jedoch in der Regel für
eine
Blütenhülle gebraucht, "die nicht in Kelch und Krone gegliedert ist" (Wagenitz 1996,
p. 278), also für eine "einheitliche Blütenhülle, bestehend aus gleichgestalteten und
gleichgefärbten Blütenhüllblättern...Das Perigon kann kronartig (Tulpe) oder
kelchähnlich sein" (Schubert/Wagner 2000, p. 413).]
"Catasetum, n., Gatt. der Orchidáceae (griech. káta=abwärts,
von...weg; lat. séta=Borste; namentl. Bezug auf die in den männlichen Blüten der meisten
Species vorhandenen zwei abwärts gerichteten Anhängsel der Säule)" (Schubert/Wagner 2000, p.147).
*Bei der Formulierung, "daß neue
Organe sehr selten oder nie plötzlich bei
einem Wesen erscheinen, als ob sie für einen besonderen Zweck geschaffen wären", setzt Darwin die
Existenz 'der Wesen' mit ihren Bauplänen und Organsystemen bereits undiskutiert voraus. Aber auch innerhalb
der Baupläne kann noch sehr viel geschehen.
**Vgl. zur 'Autoevolution' die Ausführungen von Phillip E. Johnson: "Berra's
Blunder": Der Autor bemerkt in seinem Buch DEFEATING DARWINISM BY OPENING MINDS zum Thema Learn
the difference between intelligent and unintelligent causes (InterVarsity Press,
1997, pp. 62/63):
Tim Berra is a professor of zoology at Ohio State University. He wrote a book that was
published by the Stanford
University Press with the title Evolution and the Myth of Creationism: A Basic Guide to the Facts in the
Evolution Debate. Berra's book has much the same purpose as this book. It aims to explain, for nonscientists,
how good thinkers should view the conflict between evolution and creation. Here is Berra's explanation of
"evolution," which comes illustrated with photographs of automobiles in the middle of the book:
Everything evolves, in the sense of "descent with modification," whether it be government
policy, religion,
sports cars, or organisms. The revolutionary fiberglass Corvette evolved from more mundane automotive ancestors
in 1953. Other high points in the Corvette's evolutionary refinement included the 1962 model, in which the
original 102-inch was shortened to 98 inches and the new closed-coupe Stingray model was introduced; the 1968
model, the forerunner of today's Corvette morphology, which emerged with removable roof panels; and the 1978
silver anniversary model, with fastback styling. Today's version continues the stepwise refinements that have
been accumulating since 1953. The point is that the Corvette evolved through a selection process acting on
variations that resulted in a series of transitional forms and an endpoint rather distinct from the starting
point. A similar process shapes the evolution of organisms."
Of course, every one of those Corvettes was designed by engineers. The Corvette sequence - like
the sequence of
Beethoven's symphonies or the opinions of the United States Supreme Court - does not illustrate naturalistic
evolution at all. It illustrates how intelligent designers will typically achieve their purposes by adding
variations to a basic design plan. Above all, such sequences have no tendency whatever to support the claim that
there is no need for a Creator, since blind natural forces can do the creating. On the contrary, they show that
what biologists present as proof of "evolution" or "common ancestry" is just as likely to be evidence of common
design.
I described the credentials of Professor Berra and named the publisher so nobody could accuse
me of attacking a
"straw man." A distinguished university press would not publish such a book without obtaining professional
reviews certifying that its scientific explanations were reliable. Evidently the reviewers saw nothing wrong
with equating automotive engineering and biological evolution. I am not surprised, because evolutionary
biologists typically do not understand that sequences resulting from variations on common design principles (as
in the Corvette series) point to the existence of common design, not its absence. I have encountered this
mistake so often in public debates that I have given it a nickname: "Berra's Blunder."
Nachzutragen ist jedoch, dass die Unterschiede im Organismenreich generell nicht auf
"Vervollkommnung" zurückzuführen sind, und zwar ganz im Gegensatz zu menschlichen Werken, bei denen das
häufig der Fall ist. J. von Uexküll hat diesen Punkt einmal wie folgt ausgeführt: "Man sah in der
Tierreihe den Beweis für eine stufenweise ansteigende Vervollkommnung von der einfachsten zur
mannigfaltigsten Struktur. Nur leider vergaß man dabei das eine, daß die Vollkommenheit der Struktur
gar nicht aus ihrer Mannigfaltigkeit erschlossen werden kann. Kein Mensch wird behaupten, daß ein
Panzerschiff vollkommener sei als die modernen Ruderboote der internationalen Ruderklubs. Auch würde ein
Panzerschiff bei einer Ruderregatta eine klägliche Rolle spielen. Ebenso würde ein Pferd die Rolle eines
Regenwurms nur sehr unvollkommen ausfüllen." Dasselbe gilt selbstverständlich auch für die
Botanik: Ein Alpenveilchen würde die Rolle eines Schimmelpilzes "nur sehr unvollkommen ausfüllen",
ebenso eine Pfingstrose die Rolle eines Knöllchenbakteriums.
Die Mannigfaltigkeit der Struktur, der Differenzierungsgrad der Lebensformen sagt also noch
nichts über ihre biologische Vollkommenheit im Sinne von Existenzfähigkeit aus. Andernfalls
müßten ja all die weniger differenzierten "unvollkommeneren" Formen längst ausgestorben sein; sagt
doch Darwin: "...each new form will tend...to take the place of, and finally exterminate, its less improved
parent...". - Wir haben das ja oben auch schon anhand der Beispiele von Coryanthes und Catasetum
gesehen. Ich darf noch einmal an den folgenden Punkt erinnern: "Die Coryanthes ist keine verbreitete
Orchidee und blüht außerdem in unregelmäßigen Abständen. Die Bienen sind auch nicht
gerade häufig, obwohl eine blühende Coryanthes angeblich über eine Entfernung von bis zu 8
Kilometern hinweg Bienenmännchen anzieht, werden - wenn überhaupt - nur wenige von ihnen gerade
Pollinien von einer anderen Pflanze auf dem Rücken tragen. Außerdem fällt ja auch nicht jeder
Besucher in den Eimer. Die Chancen für diese Orchidee scheinen also ausgesprochen schlecht zu
stehen. Beobachtungen der Coryanthes in freier Natur ergaben, dass die Pflanzen nur sehr selten mehr als
eine Samenkapsel pro Blütenstil trugen - in den meisten Fällen gar keine." Wie bei dieser Sachlage die
Entwicklung von Coryanthes aus einer "normalen" Orchidee durch Mutationen mit "slight or even invisible
effects on the phenotype" (Mayr) abgelaufen sein soll ("denn die natürliche Zuchtwahl wirkt nur
dadurch, dass sie sich kleine allmähliche Abänderungen zunutze macht; sie kann nie einen großen
und plötzlichen Sprung machen, sondern muss mit kurzen und sicheren, aber langsamen Schritten
voranschreiten" - vgl. Darwin oben), wobei jeder kleine Schritt mit entscheidenden Selektionsvorteilen
verbunden gewesen sein soll ("each new form will tend...to take the place of, and finally
exterminate, its less improved parent"), ist für mich nicht nachvollziehbar. Stattdessen scheitert die
Selektionstheorie an solchen Beispielen: Müßte eine solche Evolution nicht über viele
funktionsbeeinträchtigte, wenn nicht gar funktionsunfähige Zwischenglieder führen? (Siehe auch oben
die Ausführungen nach Prof. Wolfgang Kuhn.) - Ich habe
übrigens die Frage nach der "Vervollkommnung" im Organismenreich in meiner ersten Staatsexamensarbeit
(1971) (vgl. "Artbegriff den zweiten Teil
des Kapitels) und in der Augenschrift (1989) weiter diskutiert.
***Auch hier könnte man - wie zu Utricularia geschehen - den Vorschlag
unterbreiten,
erst
einmal die Genetik und Molekularbiologie von Coryanthes und Catasetum detailliert zu
studieren. Zur
Utricularia-Thematik hatte ich wie folgt geantwortet und diese Antwort trifft im Prinzip auch auf die oben
genannten Beispiele zu:
Wir wissen auf morphologisch-anatomischer und physiologischer Ebene schon sehr genau, was
erklärt werden muss. Und hier versagen bisher in sämtlichen Testdiskussionen alle mir bekannten
Evolutionstheorien völlig.
Der Vorschlag, zusätzlich zu den bekannten biologischen Daten die Genetik der
Utricularia-Falle zu studieren [oder die Genetik der Synorganisationen von Coryanthes und
Catasetum], ist
mir dennoch ausgesprochen willkommen. Die Ergebnisse dieser zukünftigen Studien werden höchstwahrscheinlich die Widerlegung der verschiedenen Evolutionstheorien auch auf dieser Ebene deutlich nachweisen und
damit die schon bekannten evolutionären Schwierigkeiten noch weiter ergänzen.
Wenn es auch wichtig ist, die Unzulänglichkeiten der heutigen Evolutionsauffassungen auch
auf der genetischen [und molekularbiologischen] Ebene weiter zu fundieren, so reichen nach allen meinen bisherigen
Erfahrungen die bekannten biologischen Daten zu Utricularia [oder Coryanthes und Catasetum
als Musterbeispiel(e) für zahlreiche ähnlich gelagerte Fälle der Synorganisation] schon
vollständig aus, um jede Evolutionstheorie in äußerste Verlegenheit zu bringen. Aber nehmen wir
an, wir hätten zusätzlich noch alle genetischen Daten - sagen wir unter anderem auch einige völlig
neue Regulator- und Targetsequenzen als Beispiele für "irreducible complexity" auf der molekulargenetischen
Ebene (so wie wir das für den Elektrorotationsmotor von E. coli und von vielen weiteren Beispielen
her kennen - ich möchte an dieser Stelle das zwar zeitaufwändige, aber doch sehr lohnende
gründlich-kritische Studium von M. Behes Darwin's Black Box empfehlen), - würde dann die Mehrheit
der zeitgenössischen Evolutionstheoretiker tatsächlich Intelligent Design als Ursache für den
Ursprung der Organismen in ihr Weltbild integrieren?
Literatur Arnold, P. (1994): Orchideen. Fotografien und Texte von Peter Arnold. Collection Rolf Heyne. Wilhelm Heyne Verlag München.
Attenborough, D. (1995): Das geheime Leben der Pflanzen. Bertelsmann Verlag. Gütersloh.
Cronin, H. (1993): The Ant and the Peacock. Cambridge University Press. Cambridge. (first published 1991,
First paperback edition 1993.)
Darwin, C. (1872/1967): The Origin of Species. Everyman's
Library. No. 811. London. Introduction by W.R.Thompson.
Und: Darwin, C. (1876): UEBER DIE ENTSTEHUNG DER ARTEN DURCH ZUCHTAUSWAHL ODER DIE
ERHALTUNG
DER BEGUENSTIGTEN
RASSEN IM KAMPFE UM'S DASEIN. VIII/592 pp. - Fussnoten. AUS DEM ENGLISCHEN UEBERSETZT VON
H.G.BRONN NACH DER 6. ENGLISCHEN AUFLAGE WIEDERHOLT DURCHGESEHN UND BERICHTIGT VON
J.V.CARUS. Stuttgart. E. Schweizerbart'sche Verlagshandlung. -
Sowie
Darwin,
C.: Die Entstehung der
Arten durch natürliche Zuchtwahl. Übersetzung von Carl
W. Neumann. Reclam. Stuttgart 1963/1981 Darwin, C. (1877): DIE VERSCHIEDENEN EINRICHTUNGEN DURCH WELCHE ORCHIDEEN VON
INSECTEN BEFRUCHTET WERDEN. In: Die Bewegungen und Lebensweise der kletternden
Pflanzen. Aus dem Englischen übersetzt von J. Victor Carus. Stuttgart. E. Schweizerbart'sche
Verlagshandlung. Darwin. Gesammelte Werke. BD.9.
Dressler, R.D. (1993): Phylogeny and Classification of the Orchid Family. Cambridge University
Press. Cambridge.
Eichelbeck, R. (1999): Das Darwin-Komplott. Riemann Verlag/C. Bertelsmann Verlag GmbH.
Ferrari, M. (1997): Tarnen, Täschen, Überleben. Die Farbenpracht im Tierreich.
Karl Müller Verlag. Erlangen.
Futuyma, D.J. (1998): Evolutionary Biology. Third Edition. Sunderland, MA. Sinauer.
Goebel, K. (1924): Die Entfaltungsbewegungen der Pflanzen und deren teleologische
Deutung, pp. 34/35. Ergänzungsband zur "Organographie der Pflanzen". Gustav Fischer Verlag, Jena.
Kuhn, O. (1951): Die Deszendenztheorie. Grundlegung der Ganzheitsbiologie.
2. Auflage, p. 149. Im Kösel-Verlag zu München. Kuhn, W. (1984): Stolpersteine des Darwinismus (Band 1),
pp. 135/136. Factum-Taschenbuch Nr. 105. - Band 2: 1985. Berneck/Schweiz. Lunau, K. (2002): Warnen, Tarnen, Täuschen. Mimikry und andere
Überlebensstrategien in der Natur. Wissenschaftliche Buchgesellschaft Darmstadt. Meeuse, B und S. Morris
(1984): Blumen-Liebe. Sexualität und Entwicklung der Pflanzen. DuMont Buchverlag Köln. Nachtwey, R. (1950): Instinkt Rätsel der Welt. 223 pp. Eberhard Brockhaus, Wiesbaden.
Portmann, A. (1970) : Im Kampf um die Auffassung vom Lebendigen. Pp. 532-548 in:
Eranos-Jahrbuch 1968: Tradition und Gegenwart. Herausgegeben von A. Portmann und R. Ritsema. Rhein-Verlag Zürich.
Sheehan, T. und M. (1994): An Illustrated Survey of Orchid Genera. Cambridge University
Press. Cambridge.
Sprunger, S. (Ed.)(1991): Orchids from the Botanical Register 1815-1847. The Illustrations
(edited by S. Sprunger in collaboration with P.J.W. Cribb and W.T. Stearn. The Texts (edited by
S. Sprunger). Birkhäuser Verlag. Basel.
Schubert, R. und G. Wagner (2000): Botanisches Wörterbuch. 12. Auflage. Verlag Eugen
Ulmer. Stuttgart.
Troll, W. (1984): Gestalt und Urbild. Böhlau Verlag. Köln Wien.
Wagenitz, G. (1996): Wörterbuch der Botanik. Gustav Fischer Verlag. Jena.
Wickler, W. (1968): Mimikry. Nachahmung und Täuschung in der Natur.
Kindler VerlagGmnH, München und Fischer Taschenbuch Verlag, Frankfurt am Main 1973.
Sie bemerken u.a. "...die Bestäuber sind Bienen der Gattung Euglossa und diese nehmen nicht 'ölige Substanzen' auf sondern Düfte."
Diese Düfte sind offenbar an "the waxy substance containing the scent" gebunden ("volatile substances").
Da Sie nun in der glücklichen Lage sind, diese Frage selbst an Ihren Coryanthes-Blüten untersuchen zu können -
beobachten Sie doch bitte einmal ganz genau, ob die Bienen vielleicht nicht auch an der wachsartigen Oberfläche
der Blüten kratzen ("Alle kratzen mit ihren bürstenähnlichen Vorderfüßen ein paar Sekunden lang
wie wild an der wachsartigen Oberfläche der Blüte") und falls das zutrifft, was sie dort sammeln.
Kann natürlich auch sein, dass sich Meeuse und Morris hier mit ihren Beobachtungen und
Interpretationen einfach nur geirrt haben.
Wie dem auch sei: Die Richtigkeit der Argumentation für intelligentes Design und gegen
den Darwinismus bleibt davon unberührt. Siehe weiter:
http://www.weloennig.de/BeautifulFactsPartI.pdf,
insbesondere auch auf der Seite 35 das längere Zitat jetzt im Zusammenhang nach dem hervorragenden
Orchideen-Fachmann Günther Gerlach ("volatile substances").
Einige Quellen, die ich jetzt (z.T. nochmal) nachgeschaut habe:
(2018) https://www.oakhillgardens.com/blog/coryanthes
(2015) Video noch mal extra: https://www.youtube.com/watch?v=_uHJGdTgtXE
(2015) https://www.youtube.com/watch?v=HD3tvVWj0QY
(2017) https://plantorchid.wordpress.com/2017/02/03/coryanthes-bucket-orchids-or-monkey-throat-orchid/
"The bees, trying to get the waxy substance containing the scent, sometimes fall to the fluid-filled bucket".
(2005) https://www.researchgate.net/publication/7896838_Juggling_with_volatiles_Exposure_of_perfumes_by_displaying_male_orchid_bees
(2003?) http://www.r-b-o.eu/RBO/html/Gerlach_G.htm
chemistry of pollinator rewards (e.g. fatty oils, floral scents)
This following page was last edited on 8 March 2021, at 18:14 (UTC).
[Weitere Quellen:]
(2018) https://floraltape.tumblr.com/post/171948338118/coryanthes-alborosea-the-white-rose-bucket
https://cedar-glade.tumblr.com/post/171931629331/coryanthes-alborosea-the-white-rose-bucket/amp
"Along with this glandular activity the main event occurs by a set of glands, called the pleurids,
that will start to drip water and coalesce with the middle section of the claw’s sites which secrete
a nice smelling oily discharge that is essentially a soup of different floral volatiles.
The glands that secretes this will continuously secrete enough to fill up the modified lip(bucket)
of the orchid below. When the lip is full with this aromatic secretion the orchid should unravel
and the male orchid bees will flock in high number to this orchid. Even after the flower
is open the pleurids will maintain their drip. Once the male bee visits this orchid it will attempt
to gather the orchids scent via an open lip location and fall into the bucket."
(1989) Volatiles: http://www.pherobase.com/database/floral/floral-taxa-species-Coryanthes-alborosea.php
(2020) https://medium.com/storybank/one-hand-washes-the-other-71e8fc6041f4 "One primary source of perfumes are
from bucket orchids(Coryanthes orchid).
They secrete an oily substance at the top of the flower which drops
into a bucket like area at the bottom of the flower. When the bees try
to get to the the top of the flower, they often fall into the bucket due to the waxy walls."
(2013) Orchids. Costa Rica (Racosi p. 59): "When the bee then visits a female flower, it again enters the lip cavity upside-down in order to gather the fragrance.
https://lagamba.univie.ac.at/fileadmin/user_upload/p_lagamba/pdf/Wiss_Infos/Orchids_of_the_Golfo_Dulce_Region.pdf
Günther Gerlach (2011, pp. 258/259): THE GENUS CORYANTHES: A PARADIGM IN ECOLOGY (LANKESTERIANA 11(3): 253—264. 2011):
"The walls of the bucket formed by the lip show cell arrangements similar to the carnivorous
pitcher plants Cephalotus follicularis Labill. (Cephalotaceae) and Darlingtonia californica Torr.
(Sarraceniaceae), which are not related to orchids (Barthlott and Ehler, 1977). The cells overlap
a bit like roof tiles, and their surfaces
are covered by epicuticular waxes (Plate 2). These waxy, shingle-like surfaces are slippery,
preventing the bees from climbing out of the bucket. The column also forms part of the trap;
the cells lie side by side and also are covered by waxes.
...Like all other Stanhopeinae, flowers of Coryanthes are pollinated by male, perfume-collecting euglossine bees.
The floral fragrance is produced by osmophores hidden under the mostly cup-shaped hypochile of the lip.
These volatile substances are both the attractant and reward for the bees.
Each bee species has species-specific fragrance preferences that may also vary geographically and seasonally.
In addition to orchid flowers, male euglossines collect volatiles from rotten wood or feces."
After collecting the fragrance compounds at the horn-shaped osmophore below the hood (hypochile)
of the lip using its feathery, foretarsal brushes, the male euglossine bee
hovers and transfers the accumulated chemicals to its hollow, inflated hind tibiae.
Available from: https://www.researchgate.net/publication/262260126_The_genus_Coryanthes_A_paradigm_in_ecology [accessed May 10 2021].
"Erläuterung zu meinem Hinweis (4. 05. 2021):
Bienen der Gattung Euglossa sezernieren Lipide welche die an der Oberfläche der
Lippe der Blüte austretenden Düfte binden / fixieren , d.h. die Biene 'spuckt' auf die Lippen-Oberfläche.
Das Bestäuber-Syndrom ist schlüssig erforscht (Thomas Eltz et al). Besten Gruß, M. B."
W-E.L. 10. Mai 2021: Wir finden also zwei unterschiedliche Quellen (siehe oben und Eltz):
Eltz et al. (2005, p. 575) (August 2005 Journal of Comparative Physiology A 191 (7):575-81)
"First, the bee lands on the fragrant surface and applies to it droplets of lipids
secreted by its labial glands in which the nonpolar volatiles are dissolved and retained,
analogous to the greasy extraction (‘enfleurage’) used in the perfume industry.
Then the bee uses setal brushes on its fore-tarsi to absorb the mixture, hovers up, and
quickly transfers the liquid to the hindtibiae by squeezing the fore-tarsal brushes
through specialized combs on the mid basitarsi. Once deposited on a hair-filled groove on
the hind tibiae, the liquid is drawn inside the cuticular pouches by what seem to be capillary forces."
Juggling with volatiles: Exposure of perfumes by displaying male orchid bees.
Available from: https://www.researchgate.net/publication/7896838_Juggling_with_volatiles_Exposure_of_perfumes_by_displaying_male_orchid_bees [accessed May 10 2021].
Eltz et al. (2007, p. 2843) (Proc. R. Soc. B (2007) 274, 2843–2848):
"To collect fragrances, the bees apply large amounts of straight-chain lipids to odoriferous surfaces
from their cephalic labial glands, which dissolve the volatiles, and the mixture is then
transferred to voluminous hind-leg pockets. Here, we show that males do in fact operate a
lipid conveyor belt to accumulate and concentrate their perfume. From the hind-leg pockets of
caged male Euglossa viridissima, deuterated derivatives of carrier lipids were consecutively sequestered,
shuttled back to the labial glands and reused on consecutive bouts of fragrance collection."
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2288683/
Ich bin Herrn M. B. dankbar für seine Hinweise zu Coryanthes. Da nun durch die weitere Forschung noch
weitere 'raffinierte' und noch komplexere, synorganisierte Details und Prozesse beschrieben wurden, ist
die Argumentation gegen die Darwinsche und neodarwinistische Deutung und die Argumentation für intelligentes Design
dieser verblüffenden Coryanthes-Phänoneme
(die nach meinen Verständnis darüber auch einen gehörigen Schuss Humor beinhalten) nur noch weiter bestätigt worden.
Jetzt zum Text von 2003: