5. DIE GEOGRAPHISCHE ISOLATION
Mit den Ausführungen von Ehrlich und Raven (1969) möchte ich nun noch einige Tatsachen aufführen, die uns zeigen, dass diese Zielsetzung für weite Bereiche des Pflanzen- und Tierreichs schon im Ansatz verfehlt ist. Wie erwähnt, waren (und sind) die Pioniere der Synthetischen Evolutionstheorie, wie die schon häufig in unserer Arbeit zitierten Autoren Dobzhansky, Mayr, Stebbins u.v.a. der Auffassung, dass die Differenzierung verschiedener Populationen durch Genfluss verhindert wird - der Genfluss ist auch das entscheidende Kriterium für den oben zitierten neodarwinistischen Artbegriff, weshalb Ehrlich und Raven die Bemühungen der neodarwinistischen Pioniere kommentieren (p. 1228): "...and they focussed their discussion of speciation on various means of interrupting that flow. In other words they emphasized the role of mechanisms isolating populations from one another."
Ehrlich und Raven gehen dann detailliert auf die Frage nach dem Genfluss in der Natur ein. Pflanzen und Tiere werden behandelt. Die Autoren weisen darauf hin, dass die Möglichkeit des Genflusses zwischen natürlichen Populationen der meisten Arten durch Besiedlung weit auseinander liegender Gebiete scharf begrenzt wird und geben dazu folgende Beispiele (pp. 1229/1230):
For example, colonies of the butterfly Euphydryas editha occur scattered throughout California, many of them separated by distances of several kilometers and some by gaps of nearly 200 kilometers. It has been demonstrated that there is almost no gene flow in this species over gaps of as little as 100 meters (27). For this reason, there seems no possibility that gene flow "holds together" its widely scattered populations. The cave-dwelling collembolan Pseudosinella hersuta occurs in a series of populations in the southeastern United States. There is no gene flow between them (28), yet they resemble one another. Clarkia rhomboidea occurs in the Great Basin of the western United States as a disjunct series of similar populations in widely separated mountain ranges. These are separated by gaps of scores of hundreds of kilometers and they are genetically highly differentiated (29). Gene flow can have no bearing on their evolution under present conditions, and we suggest that these three examples are representative of the vast majority of plant and animal distributions.
.............
There is an abundance of inferential evidence indicating that, at least in many cases, gene flow is of little or no importance in maintaining many of the phenetic units we call "species". Some of the strongest evidence, of course, comes from the wide variety of organisms with asexual reproduction. When this is obligate there is, by definition, no gene flow either within or between populations.* And yet these organisms tend to occur as phenetic species - presumably groups of individuals being kept similar by their continued existence under similar selective regimes. And, as Mayr (30) points out, the existence of groups of sibling species indicates that gene flow is not necessarily the cause of phenotypic uniformity.
*Neuere Arbeiten haben übrigens gezeigt, dass die genetische Variabilität bei asexuell sich fortpflanzenden Spezies keineswegs geringer sein muss als bei den sexuell sich fortpflanzenden (vgl. z.B. Cheliak und Dancik 1982; Futuyma et al. 1981).
Weiter ließen sich hier zahlreiche autogame Pflanzenarten und weitere Höhlenbewohner aufzählen (vgl. z.B. Culver 1982). Der letztere Autor spricht zwar von einem generellen (aber nichts beweisenden) Konsensus, dass regressive Evolution (Degeneration) bei den Höhlenbewohnern nicht einsetzen kann, solange es Genfluss mit den "Oberflächenpopulationen" gibt (p. 59), - entscheidend ist aber, dass die Höhlenpopulationen in der Regel mit den Ausgangspopulationen kreuzbar sind und fertile Nachkommen haben können. Der Fisch Astyanax mexicanus zeigt "extensive regressive evolution that happened several times independently when different surface stream populations were isolated by stream piracy" (p. 64). Trotz vielfacher Unterbrechung des Genflusses, die mit Sicherheit zumindest zwischen den verschiedenen Höhlenpopulationen gegeben ist, gehören die Ausgangs- und alle Höhlenpopulationen auch nach Auffassung der Systematiker zur selben Art.
Ehrlich und Raven versuchen in ihrem Artikel für die Gleichartigkeit der Formen trotz geographischer Isolation eine gleichartige Selektion verantwortlich zu machen. Für Höhlenbewohner entfällt hingegen der "Selektionsdruck" für optische Orientierung und das dafür vorhandene genetische Potential kann 'in Ruhe' degenerieren, d.h. durch Mutationen funktionsunfähig werden und auf diese Weise zu gleichen Formen führen. Wie jedoch ein gleichartiger "Selektionsdruck" (= differentielles Überleben) an den oft recht unterschiedlichen Biotopen mit stark differierenden Biocoenosen von Alaska, Colorado und Europa (so ein folgendes Beispiel) gleichartige Formen erzeugen soll, dürfte schon etwas schwieriger zu erklären sein (die Frage, ob die Konstanz nicht in den Formen selbst liegen könnte, wird nicht gestellt).
Im Sinne ihrer Selektionsdruck-Hypothese meinen die Autoren (p. 1230):
This is clearly what is happening in Euphydryas editha in California as well as many other butterflies with populations that are totally isolated from one another. Erebia theano populations in Alaska are only slightly differentiated from those isolated in Colorado, indeed from these in Europe. Yet we would be greatly surprised if the Colorado populations (occurring as scattered isolates) receive a gene originating in Alaska once per hundred millennia. Lycaena phaeas remains Lycaena phaeas in the Sierra Nevada of California, although almost certainly no alleles from its European or eastern American relatives have reached this area for thousands of generations. The sand crab Emerita analoga has a strongly disjunct Northern-Southern Hemisphere distribution with apparently no possibility of significant gene flow (31). In spite of this the two populations are not obviously differentiated. This is just one of many cases of a phenomenon known to marine biographers as "bipolarity" (32). Similarly, many species of plants have disjunct ranges in temperate North- and South America, with varying amounts of differentiation despite a distributional gap of thousands of kilometers (33). Another case in point is the extreme resemblance of the marine faunas of the east and west sides of the isthmus of Panama, which includes organisms to occur as pairs of relatively undifferentiated "twin species" (32). The close resemblance of the faunas remains, although the organisms on either side of the isthmus (that is, those which are restricted to warm seas) have presumably not exchanged genes for two million generations or more. Similarly, reef fishes often are remarkably similar through-out tropical seas, although gene flow among their populations is probably very reduced. The same can be said for plants on the numerous low atolls scattered through the Pacific. The plants which occur on them are identical everywhere, as contrasted with the plants on the high islands which present different selective regimes.
Schließlich stellen die Autoren fest:
Similar examples of lack of obvious differentiation in the absence of gene flow we suspect will prove to be common in all groups of organisms, just as will examples of rapid and prominent local differentiation (34) with or without gene flow.
Grant (1981, p. 56 ff.) führt weitere Arbeiten auf, die die vorliegenden Einwände von Ehrlich und Raven noch untermauern (Ehrlich und Holm 1962; Sokal 1973; Raven 1976; Levin 1979 und Ehrlich und White 1980) und kommentiert (p. 56):
These authors base their objection to the traditional viewpoint on the evidence for restricted gene flow and frequently small neighborhood sizes which we have presented in chapter 2. Certainly this body of evidence is pertinent to the question.
In reexamining the question recently (Grant 1980a), I found that neither the adherents of the traditional viewpoint nor their recent opponents had adduced sufficient evidence to support their respective claims. The traditionalists have made some plausible but glib assumptions. More hard evidence is needed than is now available to back up these assumptions. The traditional view is indeed vulnerable to attack.
Als Gegenkritik schreibt der Autor:
First, the critics consider only single-generation dispersal events. In considering gene flow on the scale of geographical races and species it is necessary to employ a larger time frame encompassing many generations.
- was mit den Ausführungen des obigen Zitats zu den Geschwisterarten, der 'Bipolarität', dem isthmus von Panama ("two million generations or more") bereits widerlegt ist.
Weiter meint Grant:
Second, the authors cited above overemphasize short-range dispersal, which is indeed the modal condition, but play down the dispersal events at the long-range end of the dispersal spectrum, which may also be biologically significant.
Das ist nun eine sehr vage Gegenkritik. Natürlich muss man, um die Dinge wieder ins Gleichgewicht zu bringen, sich zunächst verstärkt auf die Gegenbeispiele konzentrieren. Immerhin kommt der Autor zu dem Schluss, dass die Ansichten, die noch vor wenigen Jahren als sicher galten, heute nicht mehr diesen Stellenwert haben:
How, then, does the matter stand at present? The answer has to be: inconclusive. The indications point in different directions.
Briggs und Walters kommentieren die neodarwinistische Auffassung der Spezies und die Forschungsergebnisse der letzten Jahre (1984, pp. 267/268):
The species was viewed as the important evolutionary unit, gene flow amongst its members making it a breeding unit. Speciation was to be seen as the breakdown of the cohesive gene pool of the species by isolating mechanisms.
This view, that gene flow is widespread and gamodemes therefore large, has been subject to radical reappraisal in the last decade. It is now contended that gene flow in nature is much more restricted than previously thought, in the distribution of both pollen grains and seeds. Thus gamodeme sizes may be small and the 'species' may now be visualised as comprising a multitude of small gamodemes rather than a single or a few large gamodemes (Ehrlich & Raven, 1969; Levin, 1978a).
Zur Veranschaulichung seien noch einige Beispiele geographischer Verbreitung aus Nelson und Platnick (1981) zitiert (pp. 383, 378, 376):
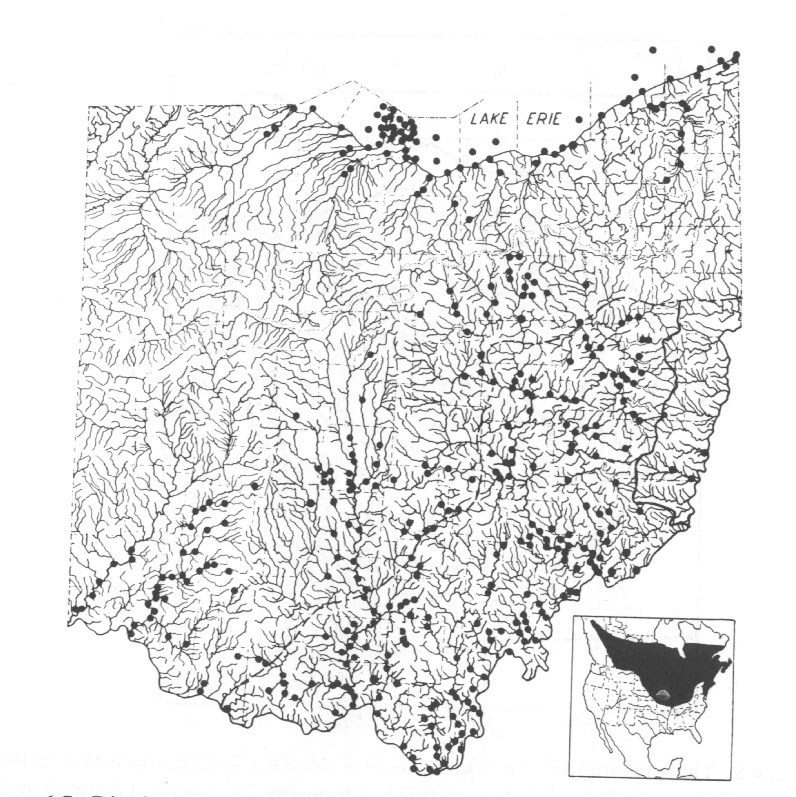
Abb. 16: Distribution of the fish, Percopsis omiscomaycus, in the State of Ohio and (inset) its distribution within North America. From M. B. Trautman (1957). Aus Nelson und Platnick 1981 nach Trautman.
Dass die verschiedenen Populationen solcher Fischarten in den verschiedenen Flusssystemen nicht durch Genfluss miteinander verbunden sind, dürfte jedem forschenden Menschen unmittelbar einleuchten. Nicht grundsätzlich anders sieht es für die Seegurke Holothuria atra aus:

Abb. 17: Distribution of the sea cucumber, Holothuria atra. Modified from Ekman (1935; see caption for figure 6.1), figure 1, p. 8. Aus Nelson und Platnick 1981.
Es dürfte klar sein, dass kein Genfluss zwischen den Populationen des Karibischen Meeres und Mikronesiens besteht.
Im Falle des Seesterns Solaster endeca sieht die Situation vielleicht etwas günstiger aus, dennoch dürfte aus der Verbreitungskarte hervorgehen, dass ein Genfluss etwa zwischen den Populationen Skandinaviens und Westkanadas und Alaskas kaum gegeben ist.
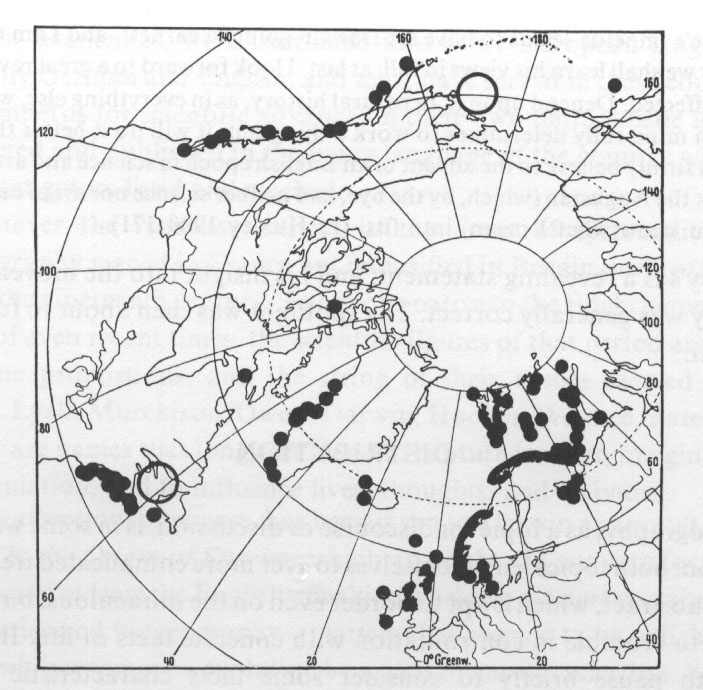
Abb. 18: Distribution of the sea star, Solaster endeca. After S. Ekman (1935), Tiergeographie des Meeres (Leipzig: Akademische Verlagsgesellschaft), figure 145, p. 257. Aus Nelson und Platnick 1981.
Es sei jedoch nicht verschwiegen, dass Nelson und Platnick für diesen Fall feststellen: "In this case, the species might be expected to occur in some at least of the intervening areas of these oceans, and in fact, its distribution might be fairly continuous throughout the oceans of the far northern hemisphere." Selbst wenn der letztere Punkt zutreffen sollte, haben wir jedoch kein kontinuierliches Verbreitungsgebiet, welches die südlicheren Populationen miteinschließen würde. (Zahlreiche weitere Beispiele für disjunkte Populationen bei Schmithüsen 1976, Walter 1979, Eigener 1982 (Verbreitungsangaben), Ehrendorfer 1983, dort weitere Literatur.)
Wie wir schon an früheren Beispielen gesehen haben, heißt Isolation nicht zwangsläufig Artbildung. Bei den oben diskutierten Beispielen von Meise, Möwe, Mensch usw. hat, genetisch betrachtet, ebenfalls keine Artbildung stattgefunden, obwohl auch hier die weit voneinander entfernt lebenden Populationen praktisch nicht mehr durch Genfluss miteinander verbunden sind.
Im Jahre 1971, p. 38 ff, habe ich die Frage unter einem anderen Aspekt behandelt. Da diese Gedankengänge die vorliegende Argumentation erweitern, möchte ich sie im folgenden mit einigen Ergänzungen wiedergeben:
Lamprecht schrieb 1966, p. 419:
Evolutionäre Schritte von niederen Kategorien zu Spezies und höheren Kategorien würden nicht zu einem natürlichen Verwandtschaftssystem, sondern zu einem Chaos führen.
...Schon Karl Ernst von Baer hatte sich ähnlich geäußert; Andermann geht auf diesen Punkt ein (1937, p.34: "Würden sich die Arten tatsächlich in einem dauernden Umwandlungs- und Vervollkommnungsprozeß befinden, so müßte man logischerweise erwarten, daß es in der Natur überhaupt keine festen Arten gäbe. Es gäbe nur ein regelloses Chaos ineinander fließender Formen und Zwischenstufen. Darauf hat namentlich K. E. von Baer mit großem Nachdruck hingewiesen. ...Es läßt sich unmöglich annehmen, daß die Entwicklung bei allen gleichzeitig lebenden Individuen einer Art gleich weit fortgeschritten ist.")
Charles Darwin greift in seiner ORIGIN OF SPECIES diese Frage auf und hat sie folgendermaßen zu lösen versucht (1859/1872, p. 172):
As natural selection acts solely by the preservation of profitable modifications, each new form will tend in a fully-stocked country to take the place of, and finally to exterminate, its own less improved parent or other less-favoured forms with which it comes into competition. Thus extinction and natural selection will, as we have seen, go hand in hand. Hence, if we look at each species as descended from some other unknown form, both the parent and all the transitional varieties will generally have been exterminated by the very process of formation and perfection of the new form.
Diese Erklärung scheint das Problem zu lösen. Sie wirkt auf den ersten Blick bestechend, logisch einwandfrei und überzeugend. Sehen wir uns den Gedanken aber etwas genauer an, so fällt folgendes auf: Als Grundlage erscheint darin die Annahme eines ständigen Vervollkommnungsprozesses ("...each new form will tend ... to take the place of ... its own less improved parent...". Von "formation and perfection of the new form", wird gesprochen), der für Darwin - genauso wie für den Neodarwinismus als Anpassungstheorie - letztlich Ausgangspunkt für die gesamte Komplexität und Mannigfaltigkeit im Bereich des Lebendigen sein soll. So logisch der Gedanke zunächst erscheint, so wenig ist er biologisch haltbar. J. von Uexküll bemerkte zu dieser Frage in seinem Buch UMWELT UND INNENWELT DER TIERE (1921, p. 3):
Man sah in der Tierreihe den Beweis für eine stufenweise ansteigende Vervollkommnung von der einfachsten zur mannigfaltigsten Struktur. Nur leider vergaß man dabei das eine, daß die Vollkommenheit der Struktur gar nicht aus ihrer Mannigfaltigkeit erschlossen werden kann. Kein Mensch wird behaupten, daß ein Panzerschiff vollkommener sei als die modernen Ruderboote der internationalen Ruderklubs. Auch würde ein Panzerschiff bei einer Ruderregatta eine klägliche Rolle spielen. Ebenso würde ein Pferd die Rolle eines Regenwurms nur sehr unvollkommen ausfüllen.
Dasselbe gilt selbstverständlich für die Botanik. Ein Alpenveilchen würde eben die Rolle eines Schimmelpilzes "nur sehr unvollkommen ausfüllen", ebenso eine Pfingstrose die Rolle eines Knöllchenbakteriums. Die Mannigfaltigkeit der Struktur und der Differenzierungsgrad der Lebensformen sagt also noch nichts über ihre biologische Vollkommenheit im Sinne von Existenzfähigkeit aus. Andernfalls müssten ja all die weniger differenzierten "unvollkommeneren" Formen längst ausgestorben sein; sagt doch Darwin: "... each new form will tend ... to take the place of, and finally esterminate, its own less improved parent ...''
Lichtig (1938) kommt unter Voraussetzung der Vervollkommnungsidee, die ja schon vorher von Lamarck publik gemacht worden ist, konsequenterweise zur Annahme einer permanenten Urzeugung; denn wie ist es anders möglich, "daß einige Lebewesen sich erst bis zur Bakterien- oder Amöbenstufe entwickelt haben, andere dagegen schon bis zur Menschenstufe, wenn doch alle zur selben Zeit als Lebewesen entstanden sind, d.h. alle aus ein und derselben Urzelle stammen und somit auch alle dieselbe Zeit zu ihrer Entwicklung zur Verfügung hatten?" Nun ist die Urzeugungshypothese schon seit Louis Pasteur (1860) widerlegt und so bleibt Lichtigs Annahme einer weltumspannenden durch alle Zeitalter bis in die Gegenwart fortdauernden Polyphylie nichts als eine Gedankenkonstruktion, die zwar - genau wie die Darwinsche - in sich logisch geschlossen erscheint, aber biologisch nicht haltbar ist, weil die Voraussetzungen nicht stimmen. Die bis heute zu findende lamarckistisch-darwinistische Idee, dass die höher differenzierten "vollkommeneren" Formen die weniger differenzierten notwendigerweise verdrängen, ist falsch. Die natürliche Selektion ist hingegen insofern ein biologisches Faktum, als dadurch weniger konkurrenzfähige Formen ausgemerzt werden können. Wovon hängt aber die Konkurrenzfähigkeit ab? Doch nicht von der Differenzierungshöhe, sondern es hägt, wie es G. E. Hiorth (1963, p. 164) für den selektiven Wert einer Mutation aufführt, von "vier Faktoren ab, der Vitalität im engeren Sinne (Wuchsgeschwindigkeit, Lebensdauer, Resistenz gegen Faktoren des nichtlebenden Milieus...), der Fertilität, der Konkurrenzfähigkeit gegen andere Genotypen oder andere Arten und der Resistenz gegen Parasiten." Vitalität im weiteren Sinne (Durchsetzungsvermögen gegenüber Faktoren des nichtlebenden und lebenden Milieus) und Fertilität haben mit der Differenzierungshöhe der Arten nichts zu tun. Die Vitalität hängt z.B. wesentlich davon ab, wie robust ein Organismus in bezug auf seine physiologische Konstitution ist. Diese physiologische Konstitution, das physiologische Gleichgewicht, wird um so anfälliger, je höher der Differenzierungsgrad eines Organismus ist, generell je feiner ein System in allen Teilen aufeinander abgestimmt und je größer die Anzahl der zu einem Ganzen integrierten Teilprozesse ist. Bergson sagte dazu treffend, dass die Entfaltung des Lebens nicht aus der Materie und ihren mechanischen Gesetzen abgeleitet werden kann. "Sie geht vielmehr gegen diese, gegen Trägheit und Zufall, zu immer höheren, gewagteren, freieren Formen." (Nach Störig 1963, p. 495.)
Und dass weiter die Fertilität nichts mit dem Differenzierungsgrad eines Organismus zu tun hat, braucht hier kaum aufgeführt zu werden. Sind doch gerade die niederen Organismen im allgemeinen fertiler als die höheren. Man vergleiche nur einmal die Fertilität von Bakterien (Generationsdauer, Zuwachsrate) oder Pilzen mit der Samenproduktion von höheren Pflanzen, Blütenpflanzen - oder bei den 'Wirbeltieren' die Fertilität des Herings mit der des Menschen!
Zum Thema Selektion ist festzustellen, dass eine wie auch immer geartete Selektion prinzipiell nichts Neues schaffen, sondern immer nur bereits Vorhandenes aussieben kann, - worauf schon die Pioniere der Genetik wie de Vries, Bateson und Johannsen mit großem Nachdruck hingewiesen haben. Künstliche Selektion führt zu Grenzen (García-Dorado/López-Fanjul 1983). Kimura hat (1968, 1983) auf biochemisch-mathematischer Ebene gezeigt, dass die Allel-Vielfalt und -Zahl die Möglichkeiten der Selektion weit übersteigt und die Theorie der 'Neutralen Evolution' ausgearbeitet. K. von Goebel begründete um die Jahrhundertwende (1898 - 1901) und in darauf folgenden Arbeiten, 'daß die Mannigfaltigkeit der Organbildung größer ist als die Mannigfaltigkeit der Lebensbedingungen'.
[...] Dass laufend durch einen Vervollkommnungsprozess gekennzeichnete "new forms" auftreten, wird einfach behauptet, das setzt Darwin einfach voraus. Aus Varietäten werden nach Darwin mit der Zeit Rassen, aus Rassen Arten, aus Arten Gattungen usw. Der Artbegriff wäre demnach fließend und Darwin hielt ihn auch ganz folgerichtig für eine reine Konvention, der keine reale Wirklichkeit in der Natur zugeordnet werden könne. Zwar ist der Neodarwinismus von diesem Artbegriff in neuerer Zeit abgerückt, aber bis heute, schreibt Lamprecht, sei der Artbegriff eine Crux für den Neodarwinismus geblieben (1966, p. 37, p. 402):
Und die Ursache ... scheint mir ausschließlich in der Annahme zu liegen, daß die Subspezies oder Varietäten im Werden begriffene Arten darstellen. Irgendwelche Beweise für die Richtigkeit dieser Annahme fehlen aber völlig. Es handelt sich nur um eine Idee.
Und davor (p. 23) zur selben Frage, ob aus Rassen Arten entstehen können:
Man meinte hiermit den Vorgang bei der Entstehung der Arten angegeben zu haben, obwohl keinerlei experimentelle Ergebnisse, geschweige denn Beweise hierfür vorgelegen haben.
Um die Diskussion noch etwas abzurunden, wollen wir nun einmal annehmen, dass alle die als hypothetisch oder falsch erwiesenen Voraussetzungen des Darwinschen Gedankenganges richtig seien. Wäre dann der schon von K. E. von Baer erhobene Einwand von dem regellosen Chaos ineinander fließender Formen und Zwischenstufen ohne jede Unterscheidungsmöglichkeit erledigt? Keineswegs! Der Vervollkommnungs- und Eliminierungsprozess würde vielleicht erklären, warum punktuell die Zwischenformen fehlen, nicht aber warum sie beispielsweise bei einer (im Extremfall sogar welt-) weiten Verbreitung von Spezies in den verschiedenen Langen- und Breitengraden von Kontinent zu Kontinent nicht auftreten. Der Vervollkommnungsprozess wäre in diesem Falle bei allen Individuen einer Art, obgleich durch schwer überschreitbare Barrieren wie Gebirge, Meere und Seen voneinander getrennt und damit befruchtungsbiologisch nicht verbunden, gleichweit fortgeschritten. Zahlreiche Beispiele liefert uns dafür die Pflanzengeographie. Ich denke hier besonders an die viel beachteten Disjunktionen von Pflanzenarten. Firbas schreibt über die arktisch-alpinen Disjunktionen (1962, p. 657):
Zahlreiche Arten der arktischen Länder findet man auch in weit südlich gelegenen Hochgebirgen, z.B. in der alpinen Stufe der Alpen, Pyrenäen, Karpaten: Salix reticulata, Dryas octopetala, Loiseleuria procumbens, Eriophorum scheuchzeri, Silene acaulis, Ranunculus glacialis u.a. Manche dieser arktisch-alpinen Arten haben außerdem noch versprengt Standorte in den höheren deutschen Mittelgebirgen, z.B. Salix herbacea, Saxifraga oppositifolia,Veronica alpina, Gnaphalium supinum; andere sogar in tiefer Lage und dann vorzugsweise in Hochmooren (Betula nana, Carex pauciflora). Die Entfernungen zwischen den einzelnen Wohngebieten betragen oft weit über 1000 km. Berühmt ist z.B. das sehr entlegene Vorkommen der rein arktischen Arten Saxifraga nivalis und Pedicularis sudetica im Riesengebirge (bei letzterer in etwa 2000 km Entfernung von ihrem arktischen Verbreitungsgebiet!). Auch die Siedlungen der in mehreren europäischen Hoch- und Mittelgebirgen auftretenden Arten sind voneinander oft weit getrennt, z.B. bei vielen Arten, die die Pyrenäen, Alpen, Karpaten, und zwar oft nur Teile dieser Gebirge, bewohnen oder die auch in manchen Mittelgebirgen (z.B. Schwarzwald, Sudeten) oder in den südeuropäischen Hochgebirgen wiederkehren (viele alpine Saxifraga-, Gentiana-, Andresace-, Soldanella-, Primula-, Potentilla-Arten u.a. ...).
Ähnliche Beispiele gibt es für europäisch-asiatisch-nordamerikanische und Steppenpflanzendisjunktionen. Wenn man auch für einige dieser Disjunktionen einen verhältnismäßig jungen Klimawechsel (Für einjährige Kräuter wären das aber immer noch Zehntausende bis Hunderttausende von Generationen. Für das Pleistozän werden 1,8 bis 2 Millionen Jahre angegeben - vgl. Krumbiegel 1981.) verantwortlich machen möchte, so ist dennoch gar nicht einzusehen, warum der Vervollkommnungsprozess von Arten wie Humulus lupulus, Anemone hepatica, Loiseleuria procumbens, Rubus chamaedorus, Phyllodoce coerulea und vielen anderen an allen Isolaten gleichweit fortgeschritten ist. Der Vervollkommnungsprozess müsste ja an allen Stellen, oft über Tausende von Kilometern hinweg, gleichzeitig und gezielt vor sich gehen. Noch komplizierter werden die Verhältnisse, wenn Firbas über die Gattungen Fagus, Acer, Aesculus und Tilia schreibt (p. 657):
Die letzteren besitzen zwar in den verschiedenen Erdteilen verschiedene Arten, diese müssen aber auf gemeinsame Vorfahren zurückgehen.
Das kann man sich natürlich vorstellen. Aber warum "müssen" diese auf gemeinsame Vorfahren im Sinne von anderen Arten mit einem anderen genetischen Potential hervorgehen? Die Entwicklung scheint ja bei den anderen oben zitierten Arten völlig stillzustehen. Warum wählt sich die Evolution nur die Gattungen Fagus, Acer und andere, um bei diesen Artbildung durch Aufspaltung zu demonstrieren, während sie Humulus, Anemone usw. völlig "links liegen" lässt, bzw. an allen Isolaten gleichzeitig in dieselbe Richtung entwickelt? Abgesehen davon handelt es sich bei den 'weiterentwickelten' Arten meist sowieso nur um Arten des morphologischen Artbegriffs; denn viele dieser Arten sind miteinander kreuzbar und können fruchtbare Nachkommen miteinander hervorbringen. Ebensogut könnte man sich doch auch vorstellen, dass Gattungen wie Fagus und Acer mit einer größeren Anzahl oder allen ihren Rekombinanten zumindest potenziell erst das ganze Verbreitungsgebiet eingenommen hat und wir heute nur noch die für das jeweilige Verbreitungsgebiet am besten geeigneten 'Arten'/Rekombinanten nach unterschiedlicher Degeneration des ursprünglich umfassenderen genetischen Potentials dort übrig geblieben sind.
In der Zoologie sieht es, was die Verbreitung der Arten betrifft, nicht anders aus als in der Botanik. Über die Verbreitung vieler Spezies schreibt Dewar in seinem Buch THE TRANSFORMIST ILLUSION (1957, p. 158):
There are hundreds of species of animals of which the geographical range is immense and in such the individuals living in Ceylon can never mate with those in the British Isles, nor can those of China or Malaya. Yet, although living in such different climates and so widely separated geographically the individuals are of the same species. The species has not split up into a number of local ones. Many species of birds have a range, which is almost cosmopolitan, such as the common kingfisher, house sparrow, osprey, sparrowhawk, merlin, kestril and barn owl. In all these the range is continuous, but the same phenomenon is seen in animals of which the range is discontinous, for example the snake Polydoniopsis melanocephalus, found in the Malay peninsula, and Archipelago, Comoro islands, Madagascar and Central America.
Der Einwand von Lamprecht und anderen - dass evolutionäre Schritte von niederen Kategorien zu Spezies usf. nicht zu einem natürlichen Verwandtschaftssystem, sondern zu einem Chaos führen, ist nach unseren bisherigen Untersuchungen voll berechtigt.