E. DEGENERATION IM ORGANISMENREICH
Statt von Degeneration spricht man in Kreisen der Synthetischen Evolutionstheorie und verwandten Auffassungen meist von "regressiver Evolution" (vgl. den Buchtitel von Schemmel et al. 1984). In einem zusammenfassenden Artikel der Naturwissenschaftlichen Rundschau (1983) wird von der "Evolution der Flugunfähigkeit bei Vögeln und Insekten" gesprochen. Mein Sprachempfinden hat mit der "Evolution der Unfähigkeit" gewisse Schwierigkeiten. Chargaff betonte 1975, p. 248:
Da das ganze Konzept der evolution aus der vor lauter Optimismus wild gewordenen VIKTORIAnischen Epoche stammt, hatte es immer ein leichtes Aroma eines überaus wünschenswerten Fortschritts. Es schien sich um die so lobenswerte Verfeinerung, Verbesserung der Welt und des Lebens zu handeln; es mußte immer nach oben gehen, und man hatte kaum von der evolution des Menschen zum Tier gesprochen, obwohl wir in unserer Zeit genug Beispiele für diesen Prozeß gehabt haben. [Großbuchstaben und kursiv vom Verfasser]
- So dass ich den auch unter Biologen bei nicht-evolutionistischen Abhandlungen allgemein respektierten Begriff der Degeneration vorziehe. Ich muss allerdings einräumen, dass der letztere Begriff nicht die faszinierenden Obertöne besitzt wie das Wort Evolution, das "Zauberwort", wie Haeckel es einst nannte, das die ganze Welt erklären sollte. Dafür ist meine Wortwahl sachlicher und dem unter dieser Überschrift behandelten Phänomenen angemessener.
Im folgenden Zitat aus dem schon erwähnten Artikel, der genauer "Degeneration der Flugfähigkeit bei Vögeln und Insekten" heißen müsste, habe ich an vier Stellen den Begriff Evolution durch Degeneration ersetzt und diese Abänderungen im Schriftbild hervorgehoben. Wie der Leser gleich feststellen wird, ergibt das in allen Fällen einen präzisen Sinn.
In dem Artikel (nach J. M. Diamond) heißt es 1983, pp. 360/361 unter anderem, dass von den 133 Rallenarten 53 flugunfähig sind und weiter:
Auch bei den Enten, Gänsen, Lappentauchern, Papageien, Ibissen, Eulenvögeln und Kormoranen kommen flugunfähige Arten vor. Die Ordnung der Laufvögel...besteht sogar ausschließlich aus flugunfähigen Vögeln.
...Vor allem auf weit vom Festland abgelegenen Inseln sind die "Nichtflieger" häufig.
...Alle Vögel sind als Junge flugunfähig. So konnte also die Degeneration zur Flugunfähigkeit sehr leicht über die Veränderung einiger weniger, die Entwicklung steuernder Gene geschehen sein.
(- Was mit Haeckel die Entwicklung auf einem frühen Stadium festschreiben heißt.) Und weiter:
Aufgrund dieses Mechanismus wäre erklärbar, warum die Degeneration zur Flugunfähigkeit offenbar so außerordentlich rasch geschieht. Das Vorkommen von flugfähigen und flugunfähigen Spezies, die nahe verwandt sind, läßt Zeiträume von weniger als tausend Jahren möglich erscheinen. Die fiugunfähige Ralle Fulica newtonii existiert in zwei Unterarten auf Mauritius und Reunion (bei Madagaskar), ebenso wie die Ralle Fulica chathamensis auf Neuseeland und den Chatham-Inseln und die Ralle Gallinula nesiotis auf Tristan und den Gough-Inseln im Südatiantik. Die beiden Unterarten existieren jeweils auf zwei entfernten Inseln. Beide haben unabhängig voneinander die Flugfähigkeit verloren und stammen höchstwahrscheinlich von je einem gemeinsamen Vorfahren ab. Noch eindrucksvoller dokumentiert die Ente Anas aucklandica die rasche Degeneration. Eine fliegende Unterart kommt auf Neuseeland und der Campbell-Insel vor, eine fiugunfähige auf der Auckland-Insel.
Und zu den flugunfähigen Insekten lesen wir unter anderem, dass sich die Degeneration zur Flugunfähigkeit hier noch rascher vollzieht als bei den Vögeln.
So ist jedes Insekt auf der arktischen Heard-Insel flugunfähig. Auf der Campbell-Insel (südlich von Neuseeland) springen "flügellose Fliegen" und Schmetterlinge wie Grashüpfer über die Insel.
Weiter heißt es, dass die Insektenpopulationen die Flugunfähigkeit "offenbar latent in ihrem Genom mit sich" trägt. Das ist sicher richtig und entspricht der allgemeinen Tendenz mutativen Informations-, Struktur- und Funktionsabbaus.
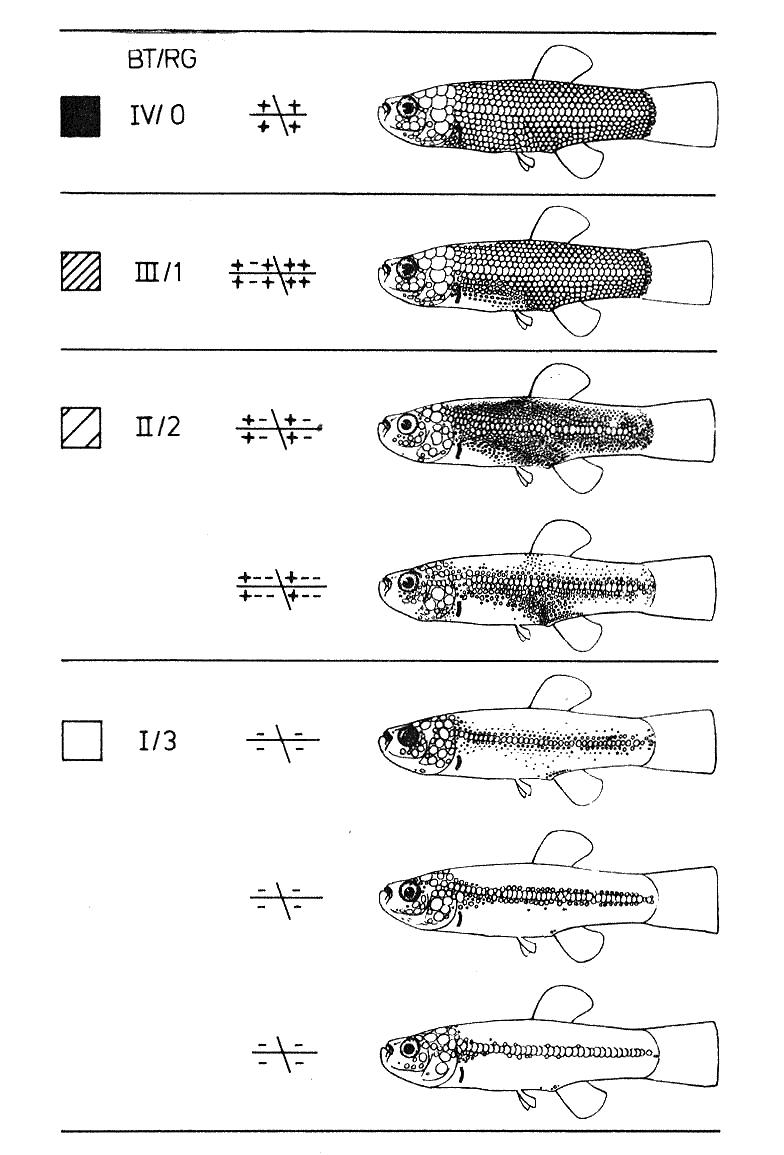
Zahlreiche weitere Beispiele führen die 8 Autoren des einleitend schon zitierten Buches "Regressive Evolution und Phylogenese" auf. Ein Beispiel sei aus Villwocks Beitrag SCHUPPEN UND VENTRALFLOSSENREDUKTIONEN, PHÄNOMENE REGRESSIVER EVOLUTION AM BEISPIEL ALTWELTLICHER ZAHNKARPFEN DER TRIBUS APHANIINI (PISCES: CYPRINODONTIDAE) zitiert. In der Abbildung werden die Regressionsgrade der Beschuppung beim Anatolichthys anatoliae-Formenkreis verdeutlicht.
Zu den Ursachen des weitverbreiteten Abbaus von Ventralflossen und Schuppen bis zum Verlust, schreibt der Autor u.a. p. 90:
Basis zum Verständnis liefert dabei die Annahme vom Fortfall der auf den Erhalt eines bestimmten Organs (allgemein: Merkmals) gerichteten stabilisierenden Selektion. Das Ergebnis ist - kurz gesagt - die Anreicherung "destruktiver" Mutanten im Pool der normale Merkmalsausbildung gewährleistenden Gene. Heterozygotie auf polygener Grundlage ist die genetische Folge. Welche Erklärung man im konkreten Einzelfall auch immer als Ausgangssituation unterstellt, das Grundphänomen rudimentierender Strukturen bleibt auf der skizzierten genetischen Grundlage stets das gleiche:
Die allen Beispielen gemeinsame, unmittelbar erkenn- oder experimentell nachweisbare hohe Variabilität während der phyletischen Übergangsphase bis zum mehr oder weniger weitgehenden Totalverlust des betreffenden Merkmals.
Die vielen interessanten Details in der gründlichen Arbeit des Verfassers kann ich hier nicht besprechen. Einen Punkt möchte ich im Rahmen dieser Arbeit aber noch hervorheben. Nach allen bislang vorliegenden Daten handelt es sich in solchen Fällen um polygen bedingte Degenerationserscheinungen. In unterschiedlichen Populationen degenerieren unterschiedliche Gene dieses Polygensystems. Wie auf den Seiten 336 u. 338 hervorgehoben, sind im Tierreich und beim Menschen dominante Gen-Funktionsstörungen wesentlich häufiger als im Pflanzenreich anzutreffen. Das trifft auch auf die Gene für die Schuppenbildung in den von Villwock aufgeführten Fällen zu (Details beim Verfasser). Daraus folgen häufig starke Interaktionen in der dann noch stärker in Mitleidenschaft gezogenen F1, was bis zur Sterilität männlicher Hybriden führen kann.
Ohne Diskussion der Frage, ob aufgrund mutativ bedingten Informationsausfalls und Strukturabbaus die in Mitleidenschaft gezogenen Fertilitätsverhältnisse in den Hybriden überhaupt für den Artbegriff relevant sein können, unterscheidet der Autor in Anwendung des neodarwinistischen Artbegriffs Geschwisterarten, wenn er z.B. p. 73 schreibt:
Im Rahmen der durchgeführten Kreuzungsanalysen hat sich ergeben, daß die zahlreichen Populationen normal beschuppter, endemisch-anatolischer Zahnkarpfen (Anatolien = asiatisches Staatsgebiet der Türkei) zwei verschiedenen Spezies, Aphanius chantrei (GAILLARD 1895) und A. anatoliae (LEIDENFROST 1942), zugerechnet werden müssen ("sibling-species") Ferner hat sich gezeigt, daß A. anatoliae Populationen graduell abgestuften, verschiedenen Fertilitätstypen angehören, wobei sich die Komplexität der Situation noch durch den Einbezug der "Anatolichthys"-Arten sensu KOSSWIG und SÖZER (1945) und AKSIRAY (1948 a, b) erhöht, deren Populationen sich in der überwiegenden Mehrzahl aus schuppenreduzierten Individuen zusammensetzen.
Vgl. dazu unsere Ausführungen zu der Kette von Schwierigkeiten, die sich für die Synthetische Evolutiontheone aus solcher Artabgrenzungsmethodik ergeben und die Argumente für den Vorschlag, den Artbegriff weiterzufassen pp. 67 - 325.
Weitere Beispiele für Degenerationserscheinungen sind bei den verschiedensten Organismen festgestellt und beschrieben worden. Im folgenden weise ich auf einige interessante Beobachtungen hin (wieder mit dem Hinweis, dass die Aufführung nur einen Bruchteil der dazu erschienenen Arbeiten zitiert).
Belford und Thompson schreiben in ihrer Arbeit SINGLE COPY DNA HOMOLOGIES IN Atriplex. I. CROSS REACTIVITY ESTIMATES AND THE ROLE OF DELETIONS IN GENOME EVOLUTION (1981):
(p. 92:) Atriplex is a cosmopolitan genus often associated with halophytic marginal environments. It has recently been shown to contain species with both C3 and C4 photosynthetic pathways (Björkman et al., 1973)
(p. 103:) Perhaps the most striking feature of the cross reactivity data is the wide range of values obtained within a group of closely related species. Reactions between Atriplex species vary about 30 per cent and 100 per cent.
(p. 104:) We...postulate that the variation in cross reactivity reflects varying amounts of deletion in the different lineages after their separation from a common ancestor. In contrast to base substitution, deletion events affect large blocks of DNA and may occur more often sporadically than gradually during evolution.
...We visualize a large ancestral Atriplex genome giving rise to lineages which have since experienced extensive deletion and decreases in net DNA content.
Die Autoren weisen wiederholt darauf hin, dass zunehmende phänotypische Spezialisierung mit abnehmendem DNA-Gehalt einhergeht. Die vier nach morphologischen und physiologischen Kriterien am stärksten abgeleiteten oder spezialisierten Arten (Atriplex rosea, A. sabulosa, A. serenana und A. fruticulosa) haben auch die kleinsten Genome, obwohl sie einen durchschnittlich etwas höheren Anteil an Single-Copy-DNA-Sequenzen aufweisen als die anderen Formen. Sie weisen auf weitere Autoren wie Sparrow et al. 1972 und Hinegardner 1976 hin, die ebenfalls zu dem Ergebnis gekommen sind, dass zunehmende phänotypische Spezialisation innerhalb verwandter Organismengruppen oft mit einer Abnahme an DNA-Gehalt verbunden ist. In ihrer zweiten Arbeit zu diesem Thema heben sie unter anderem auch die Unstimmigkeit zwischen den bisherigen Stammbäumen und den biochemischen Ergebnissen hervor (Belford und Thompson, ebenfalls 1981). Sie meinen, dass die heutigen Atriplex-Arten "during a single period of rapid speciation" entstanden seien.
Sarbhoy stellte 1980 die Verringerung des Chromatinmaterials mit zunehmender Spezialisation bei der Gattung Phaseolus (Bohne) mit ihren geschätzten 150 - 200 Arten fest.
Al-Kelidar und Richards bemerken zu ihren Studien der Gattung Delphinium (mit etwa 250 systematischen Arten; Familie Ranunculaceae) 1981, p. 632:
In diploids, there is a relationship between mean chromosome length and the amount of DNA in the nucleus, suggesting that DNA-rich areas are lost in chromosome re-organization. It is suggested that species with smaller chromosome length can be considered to be karyologically derivative.
P. von Sengbusch fasst (1979, p. 53) ein von Bachmann und Price erarbeitetes Beispiel wie folgt zusammen:
Bachmann und Price (Universität Heidelberg und Florida Technological University, Orlando) haben die Gattung der Microseridinae (amerikanische löwenzahnähnliche Compositen) näher analysiert und fanden eine drastische Abnahme des DNS-Gehaltes als Funktion der Spezialisierung. Die Ergebnisse machen deutlich, daß bei einer Spezialisierung nicht nur Ballast, sondern auch genetische Information abgeworfen wird. Die Variationsbreite vieler Merkmale sinkt dabei mit der Spezialisierung. Bei Verringerung der Genomgröße gehen proportionale Anteile der hoch repetitiven, der intermediär repetitiven und der singularen Sequenzen verloren.
Bennett führt (1985, p. 289) ein Beispiel für die Reduzierung der 1 C DNA-Menge in der Triticale-Züchtung auf:
Although this involves the loss of about 0.3 - 0.7 pg of DNA (equivalent to about 1 - 3 complete genomes in Arabidopsis thaliana) an improved phenotype, as judged by the stringent requirements of national trials in the case of 'Newton', can result.
Ähnliche Beispiele für das Verhältnis von DNA-Gehalt und Spezialisation bei Sessions et al. 1982 (Salamander), Vitturi et al. 1982 (Schnecke). Die letzteren formulieren die Ergebnisse zu diesem Punkt p. 332 folgendermaßen:
In fact, a lower number of chromosomes and a lower DNA content seem to characterize the more evolved species (...Literatur...).
In particular, this has also been obtained for certain groups of species within the phylum Mollusca (...Literatur...).
Colombera (1982) hat diese Frage an Tunikaten untersucht und ist zum gleichen Ergebnis gekommen.
Harada et al. berichten (1982), dass nach ihren Untersuchungen an japanischen Fledermäusen Heterochromatinabnahme mit 'Speziation' einhergehen kann.
Yosida hebt 1980, p. 761 in seiner Arbeit KARYOTYPE OF THE INDIAN SPINY MOUSE RESULTED FROM TANDEM FUSION OF SOME OF THE HOUSE MOUSE CHROMOSOMES u.a. hervor, dass ungefähr 13 % der Gesamt-Chromosomenlänge der Hausmaus verloren gegangen zu sein scheint, um die neue Zusammensetzung zu ermöglichen.
Green (1983) stellt die Reduktion in der Chromosomenzahl beim koreanischen Frosch Rana dybowskii auf 2n = 24 im Vergleich zur typischen Chromosomenzahl von 2n = 26 bei dieser Gattung fest (ob allerdings auch der DNA-Gehalt reduziert ist, geht aus der Arbeit nicht hervor). (Siehe auch Schmidtke et al. 1981 zur Situation beim Muntjak - Muntjacus muntjak vaginalis mit 2n = 6 Chromosomen beim weiblichen Tier und 7 beim männlichen und Muntjacus reevesi mit 2n = 46 Chromosomen - und dem Hinweis p. 192: "The longterm trends of karyotypic evolution in the phylogeny of vertebrates always involves the reduction of chromosome numbers.')
Kaul weist (1988) in seiner Monographie zum Thema MALE STERILITY IN HIGHER PLANTS (1005 pp.) darauf hin, dass genische männliche Sterilität bei den Blütenpflanzen weit verbreitet ist. Er selbst dokumentiert dieses Musterbeispiel für Genfunktionsverluste durch spontane und (seltener) durch induzierte Mutationen für "617 species and species crosses comprising 320 species, 162 genera and 43 families" (p. 15 und p. V).
Ennos berichtet 1981, pp. 127/128 über den Ausfall von Genfunktionen beim Klee u.a. folgendes:
In white clover (Trifolium repens L.) the ability to release hydrogen cyanide (HCN) from damaged leaves is under control of two unlinked genes (Corkhill 1943, Atwood and Sullivan, 1943). Locus Ac/ac determines the presence/absence of cyanogenic glucosides linamarin and lotaustralin, locus Li/li the presence/absence of their hydrolysing enzyme linamarase. Presence is dominant to absence in both cases. Both substrate and enzyme must be present for the immediate release of HCN. Thus individuals of phenotype Ac Li are cyanogenic, and phenotypes Ac li, ac Li and ac li are acyanogenic.
...cyanogenic individuals of T. repens are less heavily damaged by small herbivores than their acyanogenic counterparts (Crawford-Sidebotham, 1972; Angseesing, 1974; Dritschilo et al., 1979).
...Surveys have shown, however, that over wide geographic areas T. repens populations are polymorphic at both the loci concerned with cyanogenesis (Daday, 1954).
Sieht man sich Genotypfrequenzen an (Ac Li: 12 %, Ac li: 48 %, ac Li: 10 % und ac li: 30 %), dann ist man doch etwas überrascht, das der HCN-produzierende und damit gegen kleine Pflanzenfresser scheinbar besser geschützte Phänotyp mit nur 12 % an den untersuchten Populationen beteiligt war. So kommen denn die Autoren auch zu dem Ergebnis (p. 127):
It is concluded that the pattern of variation at Ac and Li loci cannot be adequately explained by selection which discriminates between cyanogenic and acyanogenic phenotypes.
Zera behandelt 1981 die verschiedenen Grade der Flügelreduktion bei Wasserläufern (Gerridae, Wasserschneider) und hebt u.a. folgenden Punkt hervor (p. 218):
Many species exhibit the intermediate case of various combinations of fully winged, partially winged and/or wingless morphs in the same population at the same time.
Solche Befunde sprechen stark für die Richtigkeit der Neutralen Theorie, die ja auch die Verbreitung schwach nachteiliger Gene in den Populationen postuliert und den von der Synthetischen Evolutionstheorie für so ungeheuer wichtig gehaltenen Selektionsfaktor für sekundär erachtet. Zahlreiche weitere Untersuchungen und Befunde liegen in diesem Sinne vor (z.B. Chakraborty und Fuerst 1979, Schenfield und McKechnie 1979, Latter 1981, Varvio-Aho und Pamilo 1981, Arnason 1982, Cangant et. al. 1982; weitere Beispiele bei Kimura 1983, Kahle 1984, Nei und Graur 1984).
Das heißt allerdings nicht, dass alle Ergebnisse so eindeutig und klar sind wie die oben zitierten Beispiele. Bei einigen komplexeren Fällen meinen manche Autoren, dass sowohl Selektion als auch Zufallsdrift eine Rolle spielen dürften (z.B. Nevo 1981, Nevo und Yand 1982, Nevo et al. 1982, 1983, Gilbert und Richmond 1982, Bekele 1983). Es fällt bei solchen Beispielen allerdings auf, wie schwierig es häufig ist, einen adaptiven Polymorphismus nach naturwissenschaftlichen Regeln zu beweisen, so dass die meisten Autoren für ihre Aussagen und Behauptungen den Konjunktiv wählen müssen. Zu den wenigen Ausnahmen gehört vielleicht die Arbeit von Vuorinen et al. 1981 über Temperaturadaptation gewisser Enzymformen, da sich die zur Diskussion stehenden Temperaturunterschiede auf die kinetischen Eigenschaften von Enzymen auswirken können. Auch ein Teil der oben zitierten Abbauerscheinungen könnte mit Anpassung verbunden sein (etwa der Flügelverlust von Insekten auf arktischen Inseln) - nur gehen die Polymorphismen in aller Regel weit über das hinaus, was an Anpassung notwendig ist, so dass die wenigen Prozente von Anpassungsbeispielen in keinem Verhältnis zur Gesamtvielfalt der Lebensformen stehen, und damit der größte Teil der Variabilität auch nicht mit Adaptation erklärt werden kann.
Bei all diesen Überlegungen darf man nicht aus dem Auge verlieren, dass es der genetisch und morphologisch-anatomisch redundante Teil der Lebensformen unter bestimmten Umweltverhältnissen ist, der variieren und/oder verlorengehen kann. So heben Jones et al. 1981 zu ihren Studien der Enzympolymorphismen bei Drosophila pseudoobscura u.a folgendes hervor:
More refined analyses involving improvement in electrophoretic techniques have shown that there is further genetic variation at some of the loci controlling soluble enzymes (so that, for example, the xanthine dehydrogenase locus is now known to have 27 alleles rather than the six previously detected (Singh, Lewontin and Felton 1976), while proteins that have a structural role (and that are far more abundant in the cell than are soluble enzymes) possess almost no genetic polymorphisms.
Dass alle 27 Allele einen bestimmten Selektionsvorteil haben könnten, nehmen die 5 Autoren dieser Arbeit nicht an. Nach Schätzung der Gesamtpopulation von Drosophila pseudoobscura auf 1014 (100 Billionen) Individuen und Überlegungen zur Verbreitung von Allelen meinen die Autoren p. 174:
Such large populations would allow the maintainance of many neutral alleles at a polymorphic locus without their loss to drift. The probable existence of scores of alleles at, for example the Xdh locus in this species might therefore be more easily accomodated in a neutral hypothesis or a hypothesis of weak purifying selection than was the case when the effective size of D. pseudoobscura populations appeared to be low because of restricted migrations.
________________________
ANMERKUNG ZU SEITE 408 (KLEE UND DEGENERATION VON GENFUNKTIONEN)
Die Situation erweist sich bei näherer Betrachtung als komplexer. Till kommt 1987 zu dem Ergebnis, dass verschiedene Phänotypen gleichzeitig an derselben Pflanze vorkommen können (p. 267): "That is, some leaves on a plant could be cyanogenic whereas others, on the same plant, could contain only the cyanoglucosides, only the enzyme, or neither." Sie nennt das Phänomen die "intra-individuelle Heterogenität". "The intra-individual heterogeneity means that the frequency of cyanogenesis in natural populations does not really refer to populations of plants, but rather to populations of leaves" (p. 269). Zur Eliminierung von AcLi-Genotypen bei niedrigen Temperaturen (natural rejection: vgl. p. 564) bemerkt sie p. 270: "At some locations, selection appears to have favoured plants that are capable of changing phenotype in response to some environmental cue. ...The outcome may well be the same in that the frequency of cyanogenic plant leaves in a population can change without the elimination of any individuals..." Das erinnert an die Hypothese stressinduzierter Genomveränderungen (McClintock vgl. p. 445 unten).
Zur Kälteempfindlichkeit der cyanogenen Pflanzen/Blätter: Man findet starke Schwankungen der AcLi-Frequenz bei gleicher Höhenlage und Umwelt. So gibt De Aranjo für AcLi bei 100 ft. bei einer Population 69% und bei der nächsten nur noch 32% an, bei 500 ft. 30% und 57%. Erst ab 850 ft. zeigt sich ein statistisch significanter Abfall der AcLi-Häufigkeiten. Bei 1600 ft. z.B. werden nur noch 11% festgestellt. Die erstaunlichen Differenzen bei gleicher Umwelt könnten weitgehend neutrater Natur sein! Auf der anderen Seite erscheinen mir die AcLi-Frequenzen in Abhängigkeit von Temperaturgradienten (Breiten- und Längengrade) durch die Arbeiten Dadays abgesichert zu sein: Wir haben damit ein weiteres schönes Beispiel für selektionsbedingten Genfunktionsverlust (vgl. pp. 589/590).
Die Hypothese hingegen, dass die cyanogenen Pflanzen/Blätter einen Selektionsvorteil gegen kleine (oder auch größere) Pflanzenfresser hätten, konnte in den gründlichen Untersuchungen von Bishop und Korn nicht bestätigt werden (vgl. auch p. 409). Angesichts der Ergebnisse Tills ist das wohl auch nicht zu erwarten.
Literatur:
Bishop, J.A. und M.E. Korn (1969): Natural selection and cyanogenesis in white clover, Trifolium repens. Heredity 24, 423 - 4310
Daday, H. (1954): Gene frequencies in wild populations of Trifolium repens. I. Distribution by latitude. II. Distribution by altitude. Heredity 8, 61 - 78 und 377 - 384.
De Araújo, A.M. (1976): The relationship between altitude and cyanogenesis in white clover (Trifolium repens L.). Heredity 37, 291 - 293
Till, I. (1987): Variability of expression of cyanogenesis in white clover (Trifoliom repens L.). Heredity 59, 265 - 271