D. CYTOCHROM C UND STAMMBAUMHYPOTHESEN
Wir verlassen mit diesem Thema das Gebiet der experimentell reproduzierbaren genetischen Unterschiede (und damit der experimentell-induktiven Forschungsmethodik) und begeben uns auf die Ebene vergleichender Untersuchungen, die in fast der gesamten neueren naturwissenschaftlichen Literatur von evolutionstheoretischen Deduktionen beherrscht wird. Um in alle Details zu gehen, müsste man dem Thema ein ganzes Buch widmen. Ich möchte mich hier auf einige grundsätzliche Hinweise aus der Primär- und Sekundärliteratur beschränken.
Zunächst sei hervorgehoben, dass ein Grundproblem in den meisten evolutionstheoretischen Abhandlungen völlig übergangen wird. Geissler et al. schreiben 1976, p. 630:
Über ein Drittel der Aminosäurepositionen ist in allen untersuchten Arten identisch. So wurde z.B. die Sequenz von Aminosäure 70 bis 80 überhaupt nicht verändert, und auch His 18, Cys 14 und Cys 17 sind nicht gegen andere Aminosäuren ausgetauscht worden.
...In den invarianten Regionen des Moleküls waren Austausche entweder sofort letal oder führten zu einer solchen Verschlechterung der Funktionsfähigkeit des Moleküls, daß sie sich nicht durchsetzen konnten; auch in den sich verändernden Regionen war die Austauschbarkeit meist begrenzt, da freier Austausch in der Regel zu erheblichen Veränderungen der Tertiärstruktur führt.
Über ein Drittel der Aminosäurenpositionen ist also bei den untersuchten Arten identisch. Da die Cytochrom-c-Moleküle der Wirbeltiere 104 Aminosäurenreste besitzen, sind mindestens 34 Positionen konstant. "Konstant ist weiter die Lage der Hämgruppe, die immer an den Aminosäurenresten 14 und 17 verankert ist" (Remane/ Storch/ Welsch 1980, p. 67, in Übereinstimmung mit Geissler et al. oben). Da aber auch in den veränderlichen Regionen die Austauschbarkeit meist begrenzt ist, nehmen wir für das erste funktionsfähige Cytochrom-c-Molekül eine längere spezifische Sequenz an. Wie groß ist aber nur einmal die Wahrscheinlichkeit der zufälligen Entstehung eines Polypeptids mit 34 konstanten Positionen?
Die Antwort lauted: 1 : 2034 = 1 : 171 798 000 000 000 000 000 000 000 000 000 000 000 000 000 und auf der DNA-Ebene (bei Einbeziehung von maximal 20 % neutralen Mutationen- was in Wirklichkeit zu hoch ist) 1 : 481 = 1 : 5 846 000 000 000 000 000 000 000 000 000 000 000 000 000 000 000. Mit einem Wort: nach allen vorliegenden Daten zu glauben, dass ein solch spezifisches Molekül durch Zufall entstanden ist, ist eine Glaubensinvestition mit geringer Aussicht auf Kongruenz mit der Realität. Der gezielt-intelligente Ursprung solcher Sequenzen ist wahrscheinlicher (vgl. auch p. 359)
Zeigt uns nun der variable Teil durch die Anzahl der Substitutionen den in den Lehrbüchern generell vermittelten sicheren und eindeutigen Stammbaum? Remane, Storch und Welsch geben 1980, p. 67, zu bedenken:
Die Gesamtzahl der Substitutionen allein genügt jedoch nicht, um Verwandtschaftsbeziehungen zwischen den verschiedenen Wirbeltierarten aufzuzeigen. So unterscheidet sich das Cytochrom c des Thunfisches, eines Knochenfisches, von seinen Homologen der Säugetiere nur durch größenordnungsmäßig 20 Substitutionen, während die Cytochrome des Menschen und des Pferdes bereits in 12 Aminosäuren verschieden sind.
Sehen wir uns weitere Beispiele an: Cytochrom-c-Unterschiede zwischen Mensch und Schnappschildkröte: 15 Aminosäurenreste; zwischen Mensch und Klapperschlange: 14 Aminosäurenreste; zwischen Schnappschildkröte und und Klapperschlange betragen jedoch die Unterschiede 22 Aminosäurenreste! Folglich ist der Mensch (Klasse Säugetiere) sowohl mit der Schnappschildkröte als auch mit der Klapperschlange näher verwandt, als die Schnappschildkröte mit der Klapperschlange (beides Reptilien). - Der Ochsenfrosch unterscheidet sich vom Menschen hier in 18 Aminosäuren, von der Klapperschlange unterscheidet er sich jedoch in 23 Aminosäuren und von der Schnappschildkröte in 10 Resten. Folglich ist der Ochsenfrosch (Klasse Amphibia) mit der Schnappschildkröte (Klasse Reptilia) näher verwandt als die letztere mit der Klapperschlange (auch Klasse Reptilia). Obendrein ist der Ochsenfrosch auch noch mit dem Menschen näher verwandt als mit der Klapperschlange. Von der Sonnenblume unterscheidet sich das Cytochrom c des Menschen in 42 Arninosäurenresten, vom Brotschimmel in 47. Die beiden Pflanzenformen Sonnenblume und Brotschimmel unterscheiden sich jedoch in 55 Aminosäurenresten voneinander. Fazit: der Mensch ist mit Sonnenblume und Brotschimmel näher verwandt als die beiden Pflanzenformen untereinander. Coppedge gibt 1973/1976, p. 204, folgende Zusammenfassung:
Rattlesnake cytochrome c is closer to human (14 differences) than to rhesus monkey (15), dog (21), penguin (30), or even a fellow reptile, the snapping turtle (22)! Human beings are closer to the pekin duck (11) than to a fellow mammal, the horse (12). Tomatoes and sunflowers appear closer to human than to other mammal, reptile, amphibian, fish, insect, or yeast.
Mammals are said to have evolved from reptiles; these came from amphibians, and they arose from fish. By cytochrome c comparison, however, the dog as one extreme example is closer to carp (a fish) than to rattlesnake (reptile) or bullfrog (amphibian).
The data show that in closely related organisms, the corresponding place in the chain may be occupied by amino acids which differ greatly in size, acidity, and electrical properties. Some of the differences, moreover, would have required multiple mutations in the same codon to evolve one from the other - in the face of the laws of probability.
Bei vertieften Nachforschungen dürfte noch eine ganze Reihe weiterer Schwierigkeiten zutage treten. Man müsste nämlich die Sequenzvergleiche auch auf der DNA-Ebene und zumindest auch mit anderen Cytochromen durchführen. Wie in anderen Fällen ist auch hier mit Diskrepanzen zwischen der Protein- und DNA-Ebene sowie bei den DNA-Vergleichen zu rechnen.
Fitch weist 1984, p. 240, in seinem Beitrag CLADISTIC AND OTHER METHODS: PROBLEMS, PITFALLS, AND OTHER POTENTIALS unter dem Untertitel "So many Trees and so Little Difference" darauf hin, dass die von ihm untersuchten Cytochrom-c-Sequenzen von acht verschiedenen Bakterienarten bereits 10 995 (zehntausendneunhundertfünfundneunzig!) verschiedene evolutionstheoretische Stammbäume zulassen, wobei sich die besseren von den schlechteren nur geringfügig unterscheiden:
The worst tree is only 41 replacements worse than the best one. If you know that you are distributing more than 10,000 trees into only 41 categories, it becomes a little more difficult to say that a tree close to the low end is all that much better.
Er schlägt vor, die Aminosäurensequenz-Ebene zu verlassen und sich auf die DNA-Ebene zu begeben, da hier die Zahl der evolutionstheoretisch brauchbaren Stammbäume eher überschaubar sei. Dabei ist jedoch zu beachten, dass die Stammbäume der beiden Ebenen häufig nicht kongruent sind und dass auch die "Ergebnisse" der DNA-Ebene vieldeutig sind. Wie oben erwähnt, gibt es bei dieser Frage nicht nur ein einziges Cytochrom c zu berücksichtigen, sondern mehrere verschiedene, die auf Gen-Duplikationen zurückgeführt werden (vgl. zu diesem Thema pp. 420 - 437: Evolution durch Gen-Duplikationen?).
Wu et al. bemerken 1986, p. 61, zu den Cytochrom-c-Genen DC 3 und DC 4:
DC 3 is expressed at a low level and has evolved three times faster than DC 4. [Drosophila-Gene]
Das heißt, dass sich die postulierte Entwicklungsgeschwindigkeit der verschiedenen Cytochrom-c-Gene von Gen zu Gen unterscheidet. Die Autoren meinen weiter:
This supports the neutralist view that relaxation of functional constraint is a more likely cause of accelerated evolution following gene duplication than is advantageous mutation.
Diese Aussage wird jedoch durch den Hinweis relativiert, dass sich beim Menschen einige Cytochrom-c-Gene "at an exceptionable low rate" entwickelt haben. Und das wiederum bedeutet, dass die angenommene Entwicklungsgeschwindigkeit auch innerhalb der verschiedenen Gen-Linien unterschiedlich gewesen ist.
Macht man sich nun die Vielzahl der verschiedenen Stammbaummöglichkeiten, der unterschiedlichen 'Entwicklungsgeschwindigkeiten' sowie die Differenzen zwischen Protein- und DNA-Ebene bewusst und betrachtet darüber hinaus noch die an vielen Stellen auftretenden unmöglichen Konsequenzen, die sich allein schon aus dem herkömmlichen Vergleich ergeben, so ist klar, dass die Stammbaumdarstellungen nicht den Wert haben können, den ihnen die heutigen Lehrbücher beimessen.
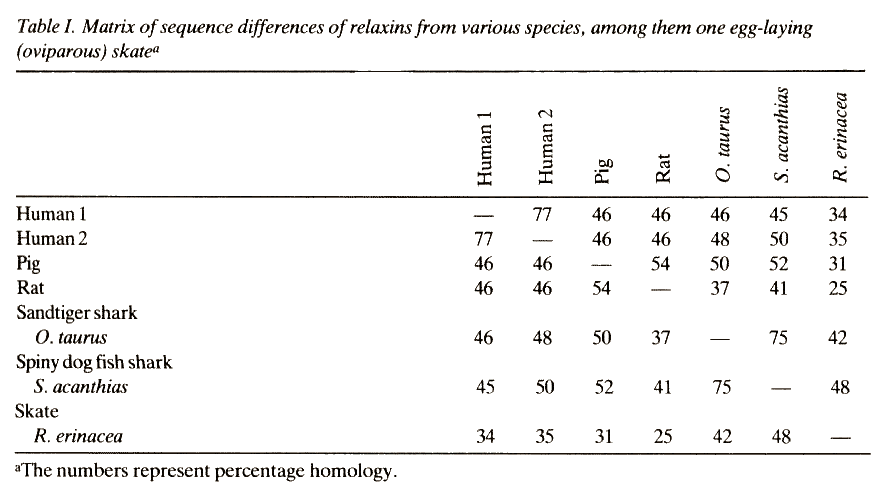
Schwabe hat 1986 weitere Kritikpunkte beim Vergleich der verschiedenen Relaxine aufgeführt. Er schreibt z.B. p. 281:
Relaxin is a hormone of vivipary in higher mammals, including man. The 55 % sequence difference between relaxins of man, pig and rat is also often explained by a rapid pace of evolution. Insulins in these species differ very little from each other because functional constraints are purportedly maintaining the structure of this hormone. By the same reasoning, rapid mutation rates are usually postulated to have occurred shortly after gene duplication and before a functional restraint had been acquired. Thus the insulin gene might have duplicated to produce the 'blanc' for a relaxin gene about 200 x 106 years ago and the extra copy might have been mutating rapidly until in placental mammals (70 x 106 years ago) a new function was fixed in position. It is troublesome, however, that shark relaxin is no more different from pig relaxin than pig relaxin is from human relaxin (see Table).
Thus, pig relaxin must still have mutated rapidly after a function had been acquired. To make matters worse, it seems quite plausible that the relaxin function might have been fixed already 500 x 106 years ago, at least among viviparous sharks. We have isolated relaxins from Squalus acanthias and Odontaspis taurus and found that these molecules cause widening of the shark birth canal analogous to the mode of action of mammalian relaxin. What seems truly astonishing is that shark relaxin also widens the pelvic bone of mice and guinea pigs; it acts specifically on structures that developed only millions of years later in different species. From this observation it must seem prudent for a molecular evolutionist to conclude that relaxin must have mutated rapidly in spite of functional constraints. Within the paradigm, however, this is considered a quirk which would best be eliminated by arguing that the receptor for relaxin leaves a much greater latitude to the hormone structure than does the receptor for insulin. This explanation loses its plausibility when one recalls that the insulin receptor can also accept variations up to 38 % of the total sequence. More amazing yet is the fact that the insulin of guinea pigs, in violation of the principles of panselectionism, 'devolved' away from the endogeneous receptor to the point where in this species the endogenous insulin is less potent than exogenous porcine insuline. What is the mechanism, we must ask, by which insulin remains nearly constant within one group of the Mammalia whereas another group uses a molecule that differs by roughly 40 % of its sequence? What mechanism is there to explain that the variation between pig and carp or pig and shark is less than the difference between the pig and hystricomorph rodents, and what allows an insulin molecule to change to a less effective state when selection is purportedly improving the fit between hormone and receptor continuously?
Der Autor weist außerdem darauf hin, dass die Relaxin-Daten nicht mit den paläontologischen Postulaten übereinstimmen: so müssen die Artentrennungen beim Relaxin B im Vergleich zum Relaxin A um etwa 350 Millionen Jahre zurückdatiert werden und weiter zweigen die Linien von Ratte, Schwein, Mensch, Hai und Rochen nach den Relaxindaten vom selben Punkt ab. Zusammenfassend stellt Schwabe 1986, p. 282 fest:
It appears that the neo-darwinian hypothesis is insufficient to explain some of the observations that were not available at the time the paradigm took shape. Three of its weaknesses discussed in this paper pertain to the fact that (1) evolutionary trees constructed from different proteins suggest the existence of different genealogies instead of a unique one, (2) genes are much older than previously assumed and (3) panselectionism is not for guinea pigs.
Wenden wir uns noch einem weiteren Punkt zur Stammbaumproblematik zu: Vielerorts kommt es auch zu starken Diskrepanzen zwischen morphologisch motivierten Abstammungshypothesen und solchen, die auf DNA-Vergleichen beruhen. Sibley und Ahlquist (1986) wollen die mit der DNA-Hybridisierungstechnik erzielten Ergebnisse für die Vögel und J. O'Brian (1986) für die Katzen zum festen Maßstab für die Verwandtschaftsverhältnisse erheben. Sie glauben, damit viele morphologisch-anatomisch begründete Zusammenhänge und Ableitungen endgültig korrigieren zu können. Die DNA-Hybridisierungsergebnisse stimmen jedoch häufig nicht mit den Proteinvergleichen überein, worauf erst kürzlich Tagle et al. hingewiesen haben. Sie stellen 1986, p. 512, bei der Diskussion der systematischen Stellung des Pandabären u.a. fest:
The most parsimonious trees constructed from the protein sequence data do not support the relationships of pandas and pinnepeds relative to other carnivores, as hypothesized from either traditional evidence or DNA hybridizations. ( Literatur- und Abbildungshinweise.)
Die hier nur erwähnten Probleme im Detail darzustellen, wäre eine umfangreiche Aufgabe für sich. Ich möchte jetzt nur feststellen, dass die für phylogenetische Ableitungen derzeit praktizierte Verabsolutierung der DNA-Hybridisierung folgende Punkte unberücksichtigt lässt: (1) Die Frage nach der Gen-Hierarchie (unterschiedliche Bedeutung von Regulator- und Strukturgenen für systematische Fragen); (2) unterschiedlicher Divergenzgrad von DNA- und Proteinsequenzen (insbesondere der weitgehend neutralen oder schwach nachteiligen Substitutionen); (3) unterschiedliche Redundanz verschiedener DNA-Abschnitte (Ohno 1985); (4) Retrovirenbefall verschiedener Organismen und dadurch bedingte Differenzen zwischen DNA-Molekülen (z.B. annähernd 1000 provirale Kopien der IAP-Elemente pro haploides Genom bei der Maus, ebenso 20-30 000 Kopien des ER-1-Elements bei derselben Spezies; vgl. An Dao Yang et al. 1986); (5) erhebliche intraspezifische Unterschiede in der DNA-Menge bei Pflanzen; vgl. p. 451); (6) mangelnde Kongruenz der phylogenetisch interpretierten Ergebnisse mit anderen molekularbiologischen Vergleichsdaten; (7) Mehrdeutigkeit vieler Ergebnisse. Schließlich sei (8) nach daran erinnert, dass nach unserer Betrachtung der verschiedenen Vergleichsmethoden auf den Seiten 188 - 192 bereits innerhalb eines Kreuzungspolygons die Ergebnisse nicht auf den gleichen Nenner zu bringen waren. Was ist dann erst beim Vergleich entfernter Gruppen zu erwarten?
Als nächstes wollen wir einen Blick auf die ohnehin seit Jahren umstrittenen Eiweißuhren werfen. Oben habe ich mit Schwabe schon erwähnt, dass unterschiedliche Proteine zu unterschiedlichen Datierungen führen können.
Andrews macht 1985, p. 498, auf folgende Probleme aufmerksam:
Two problems affect the accuracy of the protein clock. First, the fossil calibration date may be wrong because it is based an the earliest appearance in the fossil record of a species belonging to a particular lineage. It gives only a minimum date for the origin of the lineage, and the 'earliest appearance' is something that is always subject to change from new evidence. Second, the rates of change of the proteins being analysed may change along or between lineages; Goodman and others have shown that, particularly for the Hominoidea, the rate of change in a number of proteins is not constant. The Protein clock must therefore be said to be uncertainly calibrated and to keep irregular time.
Der Autor selbst glaubt, dass die Daten der DNA-Hybridisierung und -sequenzierug eine bessere Uhr liefern. Die eben zitierten Einwände zur Proteinuhr treffen jedoch prinzipiell auch auf DNA-Uhren zu und ebenso gelten die oben aufgeführten, bei phylogenetischen Interpretationen der DNA-Hybridisierung übersehenen Punkte.
Patton und Avise bemerken in ihrem Beitrag EVOLUTIONARY GENETICS OF BIRDS IV. RATES OF PROTEIN DIVERGENCE IN WATERFOWL (ANATIDAE) 1986, p. 129:
Since it now appears quite possible that homologous proteins can evolve at different rates in different phylads, molecular-based conclusions about absolute divergence time for species with a poor fossil record should remain appropriately reserved. (Vgl. auch Avise et al. 1980, Cesaroni et al. 1981, Aquadro und Avise 1982, Tagle et al. 1986.) [Siehe auch das Beispiel Seite 546 (Nachtrag).]
Kehren wir zum Cytochron c zurück: Vielleicht wird sich der eine oder andere Leser schon gefragt haben, wie sich denn aus Intelligent-Design-(ID-)biologischer Sicht die Cytochrom-c-Verhältnisse darstellen. Arbeitshypothesen:
Als erster wichtiger Punkt wäre hervorzuheben, dass das Cytochrom c nicht isoliert betrachtet werden darf, sondern im Zusammenhang mit der ganzen Atmungskette und diese wiederum mit der Gesamtorganisation der verschiedenen Lebensformen gesehen werden muss. Die Anpassungsoptima in der Funktionsfähigkeit der Atmungskette zur Umweltbewältigung werden bei Brotschimmel und Klapperschlange sowie Sonnenblume und Scbnappschildkröte sicherlich unterschiedlich sein. Entsprechend sind einige feine biochemische Unterschiede nicht nur im Cytochrom c, sondern in der gesamten Atmungskette zu erwarten, unter anderem auch im Cytochrom b, Cytochrom a und Cytochrom a3. Solche Unterschiede sollten zur möglichst optimalen Funktionsgewährleistung auch aufeinander abgestimmt sein. Von daher sind aus ID-biologischer Sicht auch größere Unterschiede zwischen stark differenten Organismen mit unterschiedlichen Anforderungen zur Umweltbewältigung als ursprüngliches Design zu erwarten. Als sicher konstatierte Naturgesetzmäßigkeit sind zusätzlich die neutralen und schwach nachteiligen Mutationen in die Betrachtung einzubeziehen. Die Organismen können von ihrem ursprünglich optimalen Leistungsniveau abdriften, aber eben dennoch weiterexistieren (s.o.). In älteren Organismen könnte sich eine größere Anzahl neutraler und schwach nachteiliger Mutationen angehäuft haben als in jüngeren Lebensformen. Weiter sind unterschiedliche Redundanzgrade bei den verschiedenen Organismen zu berücksichtigen. Bei verschiedenen Pflanzen könnten daher in den neutralen Bereichen die Unterschiede größer sein als womöglich bei Reptilien und sie wären am geringsten bei den zuletzt erschaffenen Lebensformen und ebenso bei denjenigen mit der geringsten Redundanz in den Veränderungsmöglichkeiten der Sequenzen. Das würde uns dann z.B. auch erklären, warum die Differenzen zwischen Schnappschildkröte und Klapperschlange größer sind als zwischen diesen beiden Reptilien und dem Menschen: vom gleichen oder ähnlichen Ausgangspunkt bei Reptilien driften die Sequenzen in größeren Zeiträumen durch neutrale bis schwach nachteilige Mutationen auseinander. Beim Menschen ist der ursprünglich rein funktionell bedingte Unterschied zu Reptilien noch nicht so weit abgedriftet, so dass die Unterschiede zwischen bestimmten Reptilarten größer sein können als zwischen diesen und dem Menschen.
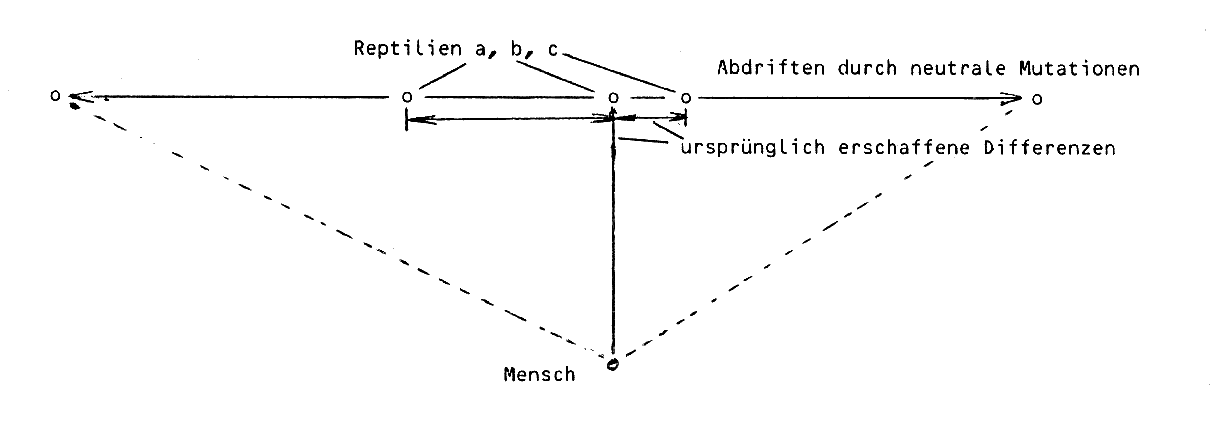
(Veranschaulichung ursprünglicher und sekundärer Unterschiede in den Aminosäurensequenzen von Cytochrom c und anderen Proteinen.)
Für diesen Ansatz spricht überdies, dass sich innerhalb bestimmter Organismengruppen die Unterschiede immer wieder in bestimmten Bereichen "clustern". Natürlich könnte ein Teil der Unterschiede auch innerhalb bestimmter Gruppen primär gegeben worden sein - etwa zwischen Brotschimmel und Sonnenblume, die ja offensichtlich für sehr verschiedene Umwelten erschaffen worden sind. Um zwischen den ursprünglich erschaffenen Differenzen und den sekundären, neutralen bis schwach nachteiligen Mutationsereignissen genauer differenzieren zu können, müssten wir einen gründlichen Aufschluss über die feineren Funktionsdifferenzen der Atmungssysteme in Relation zu den jeweiligen Organismen und ihren Umwelten haben. Solche Kenntnisse sind jedoch noch kaum erarbeitet.
Sind aber Degenerationserscheinungen im Organismenreich tatsächlich soweit verbreitet, wie unsere bisherige Argumentation nahelegt?
Mit dem nächsten Unterthema soll diese Frage weiter geklärt werden.
________________________
NACHTRÄGE ZU SEITE 402 (MOLEKULARE UHR)
Die Schwierigkeiten zur 'molekularen Uhr' haben in den letzten Jahren weiter zugenommen. Hier neuere Beispiele:
Scherer und Sontag führen (1986, p. 14/15) in ihrem Beitrag ZUR MOLEKULAREN TAXONOMIE UND EVOLUTION DER ANATIDAE (Z. f. zool. Systematik und Evolutionsforschung 24, 1 - 19) unter vielen anderen Punkten folgendes instruktive Beispiel auf:
Daß die vergleichende Interpretation von Aminosäuresequenzen nicht ohne Probleme durchgeführt werden kann, zeigt die molekulare Differenzierung von domestizierten Enten sehr eindrücklich. Während der Domestikation veränderte sich das ß-Hämoglobin der Hausente gegenüber der Wildform an 5 (!) Positionen, sodaß die Hausente, danach zu urteilen, gar nicht mehr zu den Anatidae gehört (vgl. 7). Dagegen verändert sich das [alpha]-Hämoglobin während des gleichen Zeitraums nur an zwei Positionen. Das Hämoglobin der Hausgans zeigt gegenüber der Graugans nur einen Unterschied. Ähnliche Verhältnisse können auch hei den Ovalbuminen und Lysozymen von Enten- und Hühnervögeln erwartet werden (vgl. Tab. 5, 6). Solche Befunde zeigen deutlich, daß massive mikroevolutive Änderungen von Proteinen unter geeigneten Bedingungen außerordentlich rasch ablaufen können. Der Systematiker steht vor der Frage, ob er die Sequenzunterschiede, die morphologischen, die paläontologischen oder die fortpflanzungsbiologischen Befunde stärker gewichten soll. Gehören zwei Formen zu verschiedenen Taxa, wenn sie "verschieden aussehen", mehr als einen (oder zwei?) Unterschiede in der Sequenz eines Proteins (welches?) aufweisen oder seit länger als 1 Million (?) Jahre voneinander getrennt sind? Auf dem Hintergrund von solchen Befunden und ungelösten Fragen wird besonders die Umrechnung von Aminosäuresequenzunterschieden in Jahrmillionen (für Enten vgl. z.B. OBERTHÜR und BRAUNITZER 1984; OBERTHÜR et al. 1982, 1983a, b; BRAUNITZER und OBERTHÜR 1979) massiver Kritik zugänglich. Schließlich kann niemand wissen, ob einzelne Arten nicht aufgrund funktioneller Gegebenheiten von Proteinen oder länger andauernder, extremer Isolationsphasen in kleinsten Populationen (bottle-neck-effeet, AYALA und VALENTINE 1979) einer beschleunigten Fixierung von Mutationen unterworfen waren. Wenn wir annehmen, daß die Separation von Stock- und Hausente vor 3000 Jahren (?) begann, so würde das ß-Hämoglobin eine Austauschrate von rund 1 Aminosäure pro 600 Jahren zeigen. Gewöhnlich rechnet man aber bei Hämoglobinen mit einem Aminosäureaustauseh in 6-10 Millionen Jahren (ZUCKERKANDL und PAULING 1962, vgl. OBERTHÜR et al. 1983 b), mithin hätte sich während der Domestikation die Evolutionsgeschwindigkeit des ß-Hämoglobins um dem Faktor 10000 gesteigert. Damit ist offenbar, daß die phylogenetische Interpretation von Aminosäuresequenzen per se unsinnig ist - morphologische, anatomische und vor allem paläontologische Daten müssen immer zugrunde gelegt werden.
Howgate, M.E. (1986): THE GERMAN 'OSTRICH' AND THE MOLECULAR CLOCK. Nature 324, 516:
One of the major problems with molecular clocks is that they need to be located in an absolute time frame determined by the fossil record. Undoubtedly the best such clock available at the moment is the DNA-DNA hybridization clock of Sibley and Ahlquist, but Peter Houde...argues that the DNA-DNA clock needs to be reset. ...If the picture envisaged by Houde is correct...then the datum on which DNA-DNA hybridization data is tied in to the fossil record is out by tens of millions of years.
Houde, P. (1986): OSTRICH ANCESTORS FOUND IN THE NORTHERN HEMISPHERE SUGGEST NEW HYPOTHESIS OF RATITE ORIGINS. Nature 324, 563 - 565.
P. 565: The far reaching implications of this study lie with the temporal calibration of the DNA hybridization molecular clock. Sibley and Ahlquist postulated that the divergence of ostrich DNA from rhea DNA was initiated by, and therefore could be dated by, the spreading of the Atlantic seafloor. They have since applied this calibration to a variety of other avian and mammalian taxa. With the simplicity of the vicariance biogeography hypothesis of ratite origins now challenged, one cannot accept this part of the calibration of the DNA molecular clock without at least some degree of scepticism.
Li, W.-H., M. Tanimura and P.M. Sharp (1987): AN EVALUATION OF THE MOLECULAR CLOCK HYPOTHESIS USING MAMMALIAN DNA SEQUENCES. J. Mol. Evol. 25, 330 - 342.
P. 339: The data summarized in this study clearly indicate that no global clocks apply to all mammals. In the past, rate standards have been obtained under the assumption of equal rates among mammals (Miyata et al. 1980; Li et al. 1985 a,b). It was estimated that the average synonymous rate for mammals is about 5 x 10-9 per site per year. The present study suggests that the average synonymous rate is about 6.5 x 10-9 in the rodent lineage and may be as high as 10 x 10-9 in mouse and rat, but is only 3 x 10-9 in the primate and artiodactyl lineages and may be as slow as 1 x 10-9 in humans and apes. ...nonsynonymous rate varies greatly among genes...
Fitch, W.M. (1987): COMMENTARY ON THE LI AND WU, EASTEAL LETTERS. Mol. Biol. Evol. 4, 81.
In the face of the evidence, it appears rather forced to postulate an ancient gene that is no longer with us in order to preserve the uniform-rate hypothesis. One might better conclude that the uniform-rate hypothesis is probably not reliable as a universal assumption.
Die Neodarwinisten sind stark an einer selektionistischen Erklärung von Proteindifferenzen interessiert. Entsprechend kommen aus diesem Lager 'episodic models':
Gillespie, J.H. (1986): NATURAL SELECTION AND THE MOLECULAR CLOCK. Mol. Biol. Evol. 3, 138 - 155.
P. 154: A basic conclusion of the present paper is that molecular evolution is compatible with an episodic model. To arrive at this conclusion we began with a model with varying rates of evolution. However, other starting points could lead to models without an episodic structure that are nonetheless compatible with the observed values of R (t).
Man erhält unwillkürlich den Eindruck, dass am Ende immer das herauskommt, was je nach Zielrichtung herauskommen soll. Ich, für meinen Teil, bin mehr am real-historischen Geschehen auf unserer Erde interessiert.
Immerhin weist der Autor auf folgendes Problem hin (p. 153):
Why should the rates of mutation be altered only at the time of origin of the orders of mammals and then remain unaltered for the next 60 Myr even though other lineages are branching off to form families, genera, and species? A more realistic model would have the neutral mutation rates varying through time in a single lineage to the same extent that they vary between lineages.
Schon vor einiger Zeit kamen S. Ferguson-Miller, D.L. Brautigan und E. Margoliash in ihrem Beitrag CORRELATION OF THE KINETICS OF ELECTRON TRANSFER ACTIVITY OF VARIOUS EUKARYOTIC CYTOCHROMES C WITH BINDING TO MITOCHONDRIAL CYTOCHROME C OXIDASES (1976; J. Biol. Chemistry 251, 1104 - 1115) zum Lehrbuchbeispiel Cytochrom C zu dem Schluss (p. 1104):
...the apparently constant rate at which amino acid substitutions occur in cytochrome c (13-16) were consistent with the neutral mutation hypothesis for protein evolutionary change (17-20). However, when an examination of the much more extensive data now available demonstrated that the rate of residue variation in cytochrome c was not constant, either in a single line of descent, at various evolutionary intervals or in different lines of descent during the same time, the neutral mutation hypothesis was no longer tenable for this protein (21-24).
Für unsere Arbeitshypothese p. 402 ist folgende Beobachtung der Autoren interessant (p. 1111):
Major differences were observed in the reactivity at low concentrations of four cytochromes c with cytochrome c oxidase.
...changes in primary structure have resulted in changes in the surface topography of the molecule leading to changes in binding while maintaining unaltered the essential mechanism.
Mit der Eichung molekularer Uhren durch die Fossilbefunde sind erhebliche Schwierigkeiten verbunden, die besonders in der Anthropologie in letzter Zeit zu erheblichen Kontroversen geführt haben:
Lewin, R. (1987): AFRICA: CRADLE OF MODERN HUMANS. Science 237, 1292 - 1295.
P. 1292: For many researchers...there is a head-on conflict between the molecules and the fossils. "If the molecular evidence is correct, then the fossils become inexplicable", states Milford Wolpoff of the University of Michigan. "But I believe the fossil evidence shows that the molecular biology is being wrongly interpreted."
P. 1294: Berkeley biochemists...conclusion...all modern humans derive from a population that lived about 200,000 years ago in Africa from which populations migrated to the rest of the Old World about 100,000 years later. Little or no interbreeding with existing Archaic sapiens populations occurred, suggest Wilson and his collegues. "They have calibrated the mutation rate incorrectly", claims Wolpoff. "With a much slower rate they would get a time of origin of 850,000 years ago, which I believe is correct." Smith also hesitates to accept the molecular biology evidence at face value. ...
In seinem Artikel THE UNMASKING OF MITOCHONDRIAL EVE (1987) (Science 238, 24 - 26) kommt derselbe Autor auch auf die Eichungsmethode der mtDNA-Uhr zu sprechen (p. 25):
Given a sequence divergence of 0.57% and assuming a rate of sequence divergence of 2% to 4% per million years, "the common ancestor of all surviving mtDNA types existed 140,000 to 290,000 years ago." ...Wilson [et al.]...calibrated their figure for divergence rate using the variation they measured among individuals from New Guinea, and given a date for colonization of the region of 40,000 years ago: divide the degree of variation by 40,000 and you have a figure for the amount of mutation per year.
Zu Wolpoffs Kommentar könnte man als Kontrast hinzufügen, dass man mit einer wesentlich schnelleren Mutationsrate, wie sie beispielsweise durch Transposons ausgelöst werden kann (vgl. p. 456), zu entsprechend wesentlich kürzeren Zeiträumen kommt.
Die Berechnungen von Wilson et al. beruhen auf der Besiedlung Neuguineas vor 40 000 Jahren. Wie sicher ist nun diese letztere Zeitangabe? Ein Blick in drei neuere Enzyklopädien (Britannica 1982, Universalis 1985, Meyers 1984) zeigt drei verschiedene Erstbesiedlungsdaten (über 50 000, 12 000 und über 30 000 Jahre). Eine genaue Studie der Primärliteratur dürfte die Details der Unsicherheit zutage fördern.
G. Bernardi gehört zu den führenden Molekulargenetikern, die der Neutralen Theorie und damit der molekularen Uhr kritisch bis ablehnend gegenüber eingestellt sind. P. Perrin und G. Bernardi bemerken z.B. in ihrem Beitrag DIRECTIONAL FIXATION OF MUTATIONS IN VERTEBRATE EVOLUTION (J. Mol. Evol. 26, 301 - 310; 1988) p. 310:
...the directionality of the compositional changes and their evolutionary conservation strongly point to compositional constraints on the genome and on its compartments in vertebrates.
...these constraints do not appear to be compatible with the neutral theory of evolution (Kimura 1968, 1983, 1986). Indeed, if most third position changes were neutral, as claimed, one would not expect compositional constraints which depend upon environmental factors.
Die Deutung des G-C shifts in den 3. Codon-Positionen als Temperatur-Adaptation (beim Vergleich von Säugetieren mit 'Kaltblütern') hat jedoch auch ihre Schwierigkeiten: Er findet nämlich die gleiche Tendenz bei monokotylen PfLanzen im Vergleich zu dikotylen. Beide Gruppen kommen jedoch weltweit unter allen nur denkbaren Klima- und Temperaturbedingungen gleichermaßen vor.
Auf der Proteinebene arbeiten selbst die entschlossensten Selektionisten zur Zeit mit nur maximal 15 % der untersuchten variierenden Loci als mögliche Anpassungserscheinungen. (Vgl. z.B. R.S. Singh und L.R. Rhomberg 1987: A COMPREHENSIVE STUDY OF GENIC VARIATION IN NATURAL POPULATIONS OF DROSOPHILA MELANOGASTER: II. ESTIMATES OF HETEROZYGOSITY AND PATTERN OF GEOGRAPHIC DIFFERENTIATION. Genetics 117, 255 - 271.) Unter solchen Voraussetzungen wäre es höchst erstaunlich, wenn der DNA-Code vollständig, d.h. selbst noch an den vierfach degenerierten 3. Codon-Positionen, funktional determiniert wäre. Wie erklärt sich dann die größere Variabilität in der 3. Position im Vergleich zu der 1. und 2.? Meiner Auffassung nach reichen weder Zufalls-Substitutionen noch Adaptationen für eine vollständige Erklärung von "Linear G" (vgl. p. 563) - obwohl beides vorkommt. Dabei sind noch die genauen Anteile zu bestimmen. Es ist das Verdienst Bernardis, mit gut fundierten Ergebnissen die totalitäre Zufallsauffassung und damit die molekulare Uhr in Frage gestellt zu haben.
Patterson kommentiert den Fragenkomplex 1987, p. 17, wie folgt:
Since it was first proposed in the mid-1960s, the clock hypothesis has been one of the most controversial topics in evolutionary theory.
Und er weist auf mehrere Autoren des von ihm herausgegebenen Buches hin, die diese Uhr ablehnen (Andrews, Goodman, Miyamoto und Czelusniak) (C. Patterson 1987: MOLECULES AND MORPHOLOGY IN EVOLUTION: CONFLICT AND COMPROMISE. Cambridge.)
Die Palette der verschiedenen Auffassungen zur molekularen Uhr zeigt meiner Auffassung nach mit hinreichender Deutlichkeit die derzeitige Unsicherheit dieses Ansatzes zur Frage nach sicheren Zeitangaben!
Selbst wenn man einmal unterstellen würde, dass die molekulare Uhr gleich einem Ouarzwerk regelmäßig 'ticken' würde ( - wie ich schon betont habe, stelle ich nicht generell die Anreicherung von neutralen bis schwach nachteiligen Mutationen in den verschiedensten Populationen in Frage, sondern nur deren postuliertes, absolut regelmäßiges Auftreten zur absoluten Zeitmessung über größere Zeiträume - ), so müssten zu einer Zeitableitung aufgrund von DNA-Unterschieden über die in den meisten Arbeiten üblichen Parameter hinaus noch folgende weitere Faktoren beachtet werden ( - wobei der 3. Punkt auch schon wieder die postulierte Regelmäßigkeit in Frage stellt):
1. Die Zahl (Mittelwerte) der Replikationen von einer Zygote bis zur nächsten Generation (genauer bis zur Gameten- und Zygotenbildung an oder in der Parentalgeneration zur Erzeugung der Tochtergeneration). - Neuere Lehrbücher wie Knilppers 1985, Lewin 1987 u.a. geben 1 Austausch pro 10-9 - 10-10 Nukleotidpaare pro Replikation (!) an. Diese Replikationszahlen unterscheiden sich mit Sicherheit z.B. beim Vergleich von 1-, 2- und mehrjährigen Pflanzen und ebenso bei verschiedenen Tiergruppen. Aber auch innerhalb einjähriger Pflanzen dürften erhebliche Unterschiede zwischen größeren Pflanzengruppen in Abhängigkeit von der Zahl der Mitosen bis zur Zygoten- (und anschließenden Samen-) Produktion bestehen. Die Unterschiede in der Wachstumsgeschwindigkeit, in den Größen (Lägen/Höhen) und in den Blühterminen sind unübersehbar. Unterschiede in der Zahl und Teilungshäufigkeit der Initialzellen lassen eine Variation von 100 Prozent und mehr möglich erscheinen (genaue Daten sind mir zum letzteren Punkt bisher nicht bekannt - für Hinweise wäre ich dankbar).
2. Daraus folgt die Bedeutung des Fortpflanzungsalters, die Länge der Fortpflanzungsperiode und für viele Pflanzen- und Tiergruppen die Frage nach dem Lebensalter überhaupt. Die Generationszeit wird neuerdings bei Tieren zwecks Eliminierung von Widersprüchen molekularer Zeitangaben zunehmend berücksichtigt (vgl. z.B. W.-H. Li und M. Tanimura 1987: The molecular clock runs more slowly in man than in apes and monkeys; Nature 326, 93 - 96. P. Foley 1987: Molecular clock rates at loci under stabilizing selection; Proc. Natl. Acad. Sci. 84, 7996 - 8000).
3. Die Zahl der Nachkommenschaft. Sie variiert bei Pflanzen und Tieren ganz erheblich. Dass ein Elephant weniger Nachkommen erzeugt als ein Hering, ist allgemein bekannt (vgl. auch pp. 465/466). Ein Beispiel für die Angiospermen: Hordeum vulgare (Sommergerste): Verhältnis von Aussaat zu Ertrag etwa 1 : 40 (Produktion pro Halm etwa 20 - 45 Samen); Pisum abyssinicum 26 - 40 Samen, Antirrhinum majus 1000 - 10 000 Samen (je nach Umweltverhältnissen und Zahl der Bereicherungstriebe); Verbascum Thapsus um 700 000 Samen. Nach dem Gesetz der Rekurrenten Variation müssen bei 17 000fach höherer Samenproduktion pro Pfanze (Beispiel Pisum-Verbascum) bestimmte Mutanten entsprechend häufiger auftreten. Wenn auch die Selektionstheorie nur zum Teil mit der Realität kongruent ist, so sollten doch höhere Nachkommenschaften mit größerer Anpassungfähigkeit im Rahmen der mutationsgenetischen Möglichkeiten einhergehen (Einschränkungen vgl. pp. 466, 467, 586, 587; vgl. auch J.A. Endler 1986: Natural selection in the wild; Princeton, New Jersey). Sowohl bei Anpassungs- als auch Degenerationsprozessen könnte die molekulare Uhr durch die unterschiedlichen Zahlen der Nachkommenschaft beeinflusst werden.
Weiter dürfte hier die Poputationsstruktur eine Rolle spielen: Bei Arten mit zahlreichen kleinen diskontinuierlich verteilten Populationen können sich Mutanten eher durchsetzen als bei solchen mit weiten kontinuierlichen Populationen (Bottleneck-Frage, 'Zufallsdrift'). Weiter muss mit Unterschieden im Genom der Arten selbst gerechnet werden: Transposonfrequenzen, repetitive Sequenzen, unequal exchanges etc. sind nicht bei allen Arten gleich. ["Like selection, molecular drive can both retard and accelerate the rate of evolution in a given region of DNA. As with real clocks, the measurement of time can be affected by both internal and external interference. It is unlikely that observed rates of DNA change represent the monotonous measure of molecular metronomes." G.A. Dover (1986): Molecular drive in multigene families: how biological novelties arise, spread and are assimilated. TIG 2, 159 - 165.] Dazu kommt ein unterschiedliches genetisches Anpassungspotential (p. 587). D.h. während bei einigen Arten bestimmte Gene unter starkem 'Selektionsdruck' stehen (natural rejection bei Mutationen), können sie bei anderen Formen in Ruhe degenerieren.
Schließlich müssen noch mutagene Umweltfaktoren berücksichtigt werden (Temperaturunterschiede, Virusinfektionen, unterschiedliche Strahlungsintensitäten, Klimaschwankungen und Katastrophen globalen Ausmaßes im Laufe der Erdgeschichte etc. [letzteres Thema war lange Zeit von der aktualistischen Geologie mit einem Denkverbot belegt worden, inzwischen werden solche Fragen jedoch lebhaft diskutiert (vgl. z.B. Science 239, 977 (1988) und Scientific American 258, 24 - 25 (1988), wo die seit einigen Jahren leidenschaftlich diskutierte Impact-Frage weiter im Brennpunkt steht) - der Umschwung ist seit etwa 1980 zu beobachen]. Wie unter all diesen variierenden Parametern eine molekulare Uhr im neutralen Bereich auch nur über kleinere Zeitperioden geschweige denn über die gesamte Erdgeschichte bei allen Arten und Lebensformen absolut konstant geblieben sein soll, erscheint mir unerfindlich.
Bei all diesen Fragen erhebt sich im Hintergrund noch ein Problem, das noch kurz folgendermaßen umrissen sei:
Nach herrschender Auffassung ist die Entfaltung aller morphologisch-systematischen Unterschiede völlig unabhängig von den 'House-keeping Genes' abgelaufen. Stammbäume sind nur möglich aufgrund der unbewiesenen Voraussetzung monophyletischer Abstammung und gleichlaufender molekularer Uhren. Die Stammbäume selbst können die monophyletische Evolution nicht beweisen. Unter den Voraussetzungen der Polyphylie und Nicht-Konstanz der Nukleotid-Substitutionen auch in der 3.Codon-Position ('vierfach degeneriert') wird man die Sequenzdifferenzen entsprechend unterschiedlich interpretieren.