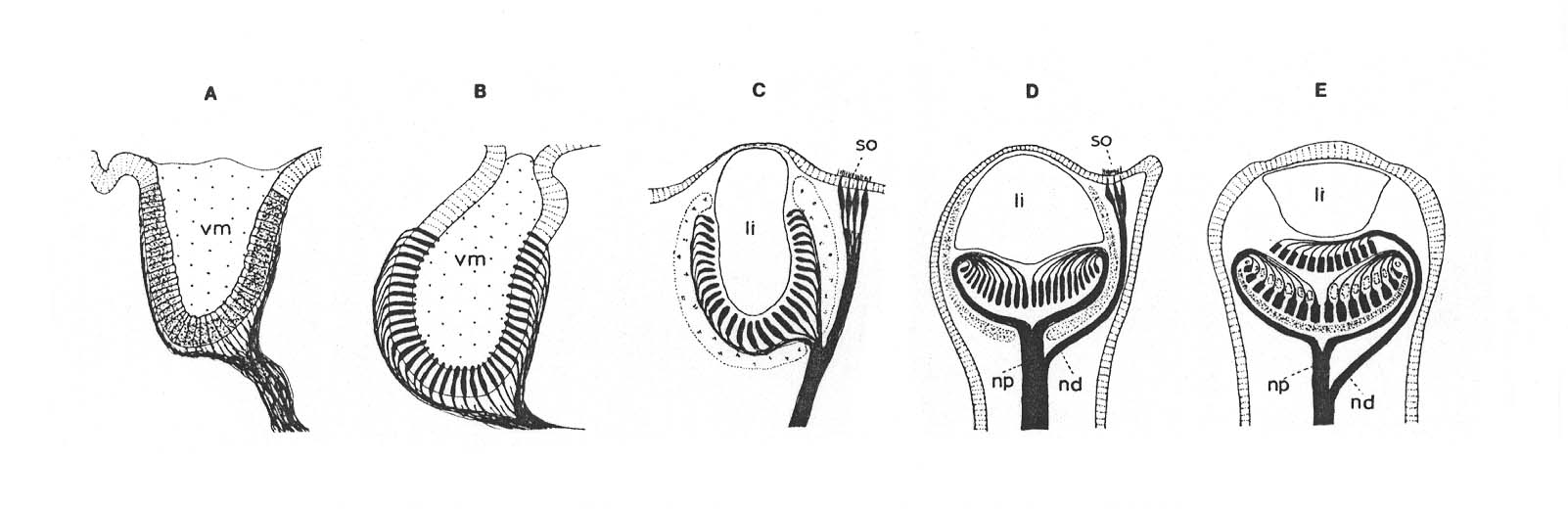
DIE MORPHOLOGISCHEN SERIEN
Nicht ohne Grund haben Salvini-Plawen und Mayr geschrieben, dass bei lebenden Tieren "an almost complete gradation from exceedingly simple light-sensitive receptors to highly complex eyes" existiert. Sieht man sich die Serie näher an, so stellt man mehrere Diskontinuitäten fest.
I. Morphologische Serie bei den Bivalvia
Morphological sequence of differentiation (including inversion) in eyes at the mantle edge of Bivalvia (cf. Hesse, 1908; Küpfer 1916; Stasek 1966; Barber et al.,1967; and others): A, everse eye pit of Arca barbata (Arcacea); B, everse pinhole eye of Lima squamosa (Pectenacea); C, everse closed lens eye and ciliary sense organ of Tridacna maxima (Cardiacea); D, closed lens eye with inverted retina cells, and ciliary sense organ in Cardium muticum (Cardiacea); E, closed lens eye or Pecten maximus (Pectanacea) with proximal retina as in Cardium, but with inverted ciliary sense organ representing the distal retina. (li, multicellular lens; nd, distal branch of optic nerve; np, proximal branch of optic nerve; so, ciliary sense organ; vm, vitreous mass.)
Abb. aus Salvini-Plawen und Mayr, p. 229
Die Autoren bemerken zu dieser Serie u.a. (p. 238):
The sequence from open epidermal photoreceptors (Arca) to almost closed everted eyes (Lima = Pectenacea) and to the closed eyes with a multicellular lens (Tridacna; Stasek, 1966) offers no problem.
Aus naturwissenschaftlicher Sicht muss man jedoch auf folgende offenen Fragen und Probleme hinweisen:
1.
Es liegt hier keine Abstammungslinie vor. Die Serie besteht stattdessen aus Vertretern dreier verschiedener Überfamilien der Muscheln (Bivalvia):
Arcacea (Archenmuscheln)
Pectenacea (Kammmuscheln)
Cardiacea (Herzmuscheln)
Mit anderen Worten: Das everse Grubenauge von Arca barbata (Arcacea) hat sich nicht zum ebenfalls eversen Lochkameraauge von Lima squamosa (Pectenacea) und dieses nicht zum Linsenauge von Tridacna maxima (Cardiacea) weiterentwickelt.
Die folgenden systematischen und paläontologischen Tatsachen dürften diesen Punkt noch weiter untermauern:
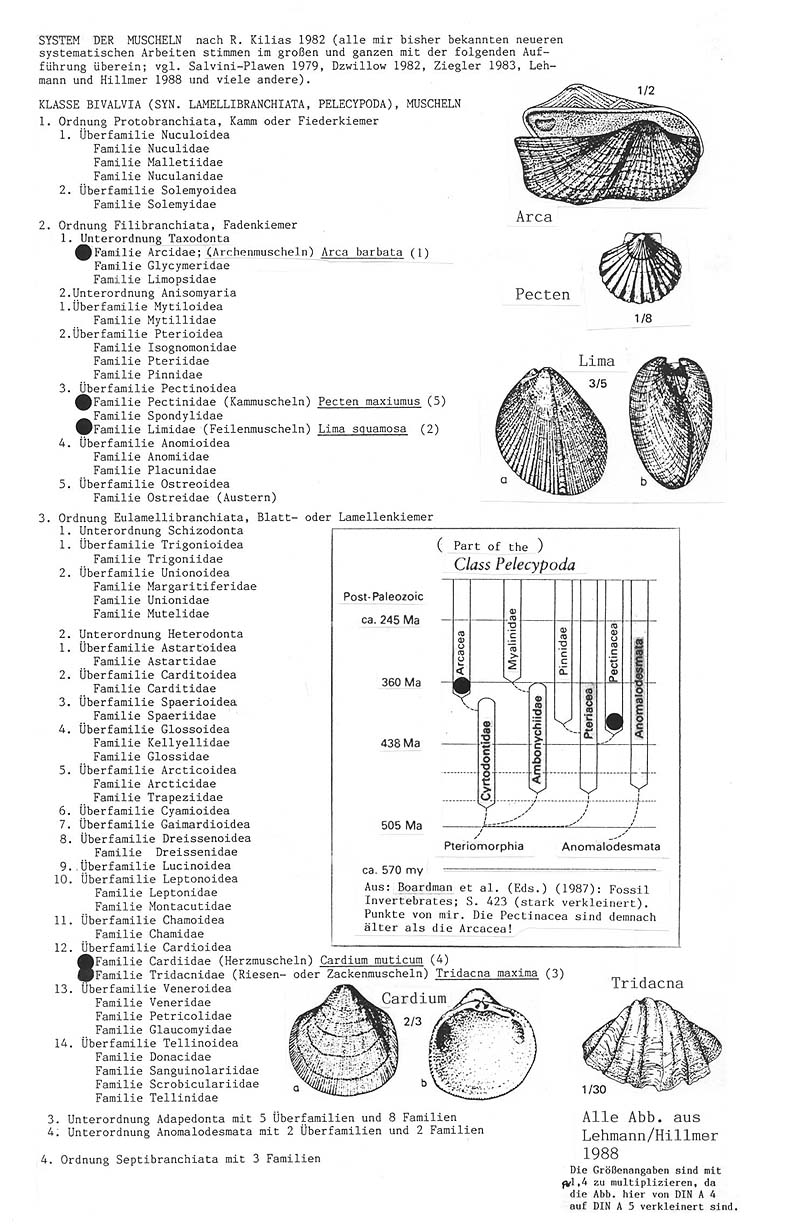
Wie der Leser sofort erkennen kann, verteilen sich hier die von Salvini-Plawen und Mayr in einer morphologischen Serie angeordneten Gattungen auf 5 Familien und 2 Ordnungen. Auch nach abstammungstheoretischen Voraussetzungen kann es sich damit nicht um eine Abstammungslinie handeln, sondern nur um die Zweigspitzen des Stammbaums.
Der angenommene Stammbaum wiederum stimmt nicht mit den paläontologischen Befunden überein, wie die folgende Abbildung aus dem Lehrbuch der Paläozoologie von A.H. Müller verdeutlicht:
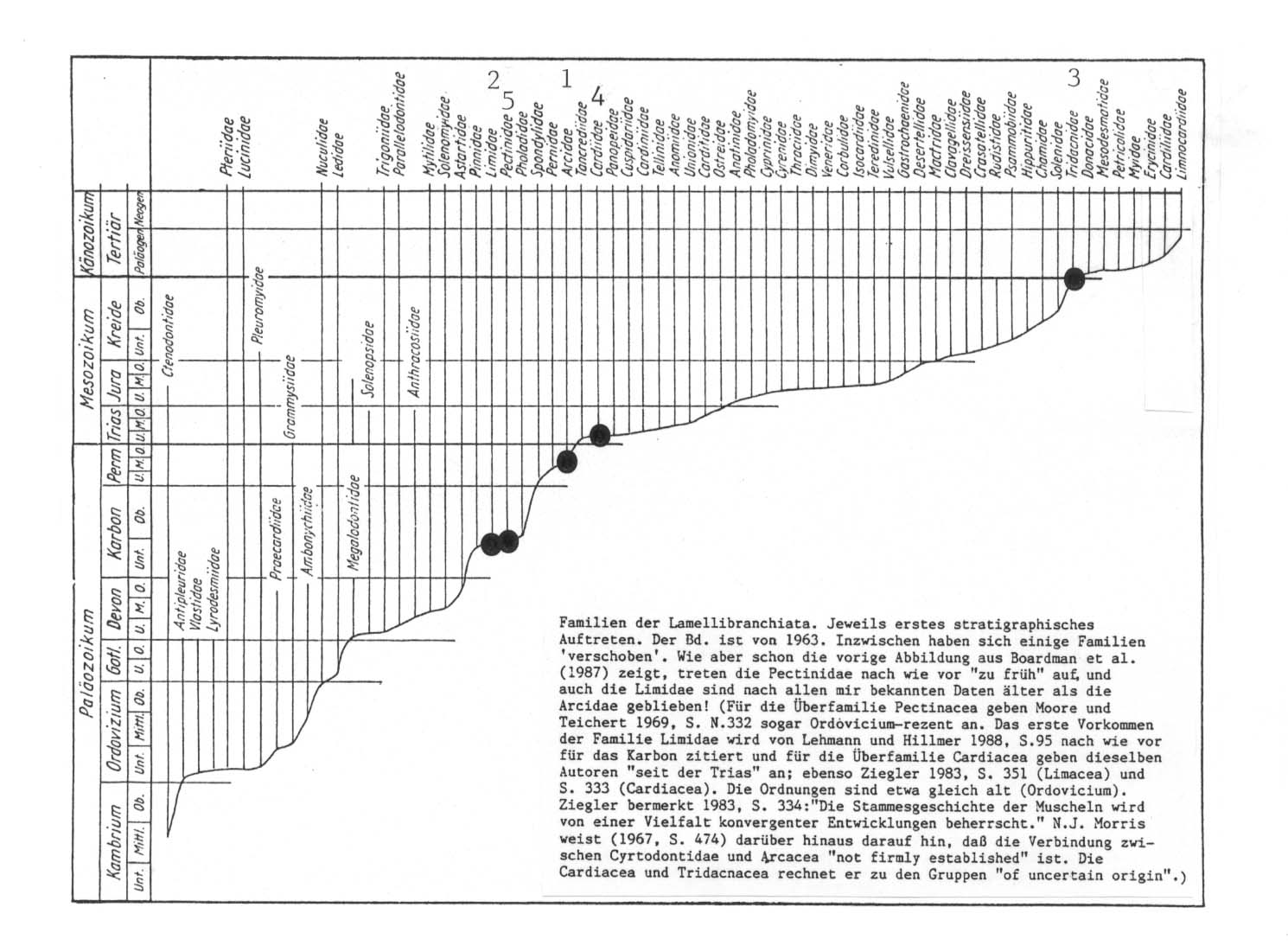
Die Reihenfolge der Gattungen in der morphologischen Serie stimmt weder mit der phylogenetisch gedachten Systematik überein (vgl. die Zahlen hinter den Gattungen), noch findet sich eine zeitliche Bestätigung des Auftretens der verschiedenen Familien durch die Paläontologie (vgl. wiederum die Zahlen über den Familien): Limidae und Pectinidae sind nach dem heutigen paläontologischen Kenntnisstand älter als die Arcidae. Tridacnidae und Cardiidae sind jünger als die Pectinidae und können folglich nicht die Ausgangspopulation für die Letzteren geliefert haben.
Salvini-Plawen und Mayr schreiben 1977, pp. 237/238:
Many Bivalvia posses photoreceptors at the mantle edge, some of which are everse pits (Arcidae, Limidae), others are everse (Tridacna) or inverse (Cardium) closed eyes with lens, or even provided with a second (distal) layer of inverse retinal cells (Pectinidae). We consider all these epidermal formations to be homologous according to the schematic sequence of levels of differentiation shown in Fig. 8 (p. 229). (Vgl. die Abb. p. 90)
Da nach dem gegenwärtigen systematischen und paläontologischen Kenntnisstand die Formen der morphologischen Serie nicht voneinander abstammen können, müssten sich nach abstammungstheoretischen Voraussetzungen auch verschiedene Differenzierungsstufen der Lichtsinnesorgane bei mehreren Familien der Bivalvia unabhängig voneinander herausgebildet haben (Konvergenz statt Homologie)*.
Man muss dabei noch berücksichtigen, dass nach Kilias "Augen nur bei einigen Muschelgruppen auftreten" (1982, p. 157; ähnlich Land 1981, p. 506, Tabelle).
*Nach den Homologiekriterien kann man auch den Homologiebegriff hier beibehalten. Homologie ist dann nicht Abstammung!
2.
Die Schritte von den angenommenen (aber hier nicht dargestellten) Vorstufen bis zu A sowie die Schritte von A nach B sind zwar auf dem Papier leicht machbar, erscheinen jedoch bei näherer Betrachtung etwas unwahrscheinlicher als die Zeichnung und die Theorie vermuten lassen. Nehmen wir nur einmal an, wir selbst sollten zwecks Vervollkommnung unserer Ausrüstung mit Lichtsinnesorganen ein zusätzliches Paar kleiner Augen bilden, sagen wir am linken und rechten Arm oberhalb der Handgelenke (innen); - oder vielleicht noch besser auf den Handrücken. (Das wäre doch eine ausgesprochene "Vervollkommnung": Man könnte gleichzeitig nach vorn und nach beiden Seiten und mit leichter Drehung auch nach hinten blicken, was eine gewaltige Erweiterung des Blickfeldes bedeuten würde.) Und jetzt wollen wir uns auf die Suche nach Kandidaten unter den über den 6 Milliarden Menschen begeben, die
a)
erst verstreut und dann auf einer begrenzten Fläche des Handrückens Lichtrezeptoren aufzuweisen haben.
b)
die mit der Zeit eine kleine erbliche Vertiefung auf dem Handrücken zeigen, und zwar an der Stelle der maximalen Dichte der Lichtrezeptoren.
c)
die in nachfolgenden Generationen über Tausende von Mikromutationen die Vertiefung an eben derselben Stelle erblich bis zum Lochkameraauge vervollkommnen (später unter Produktion eines gelartigen Glaskörpers, Augendeckels und Schutzwalls rundherum).
Wir brauchen an dieser Stelle nur wieder auf die eingangs zitierten Unwahrscheinlichkeiten hinzuweisen (vgl. p. 5 - 22), um die praktische Unmöglichkeit eines solchen Zufallsprozesses klar zu machen: Hier helfen weder große Zeiträume noch die schärfste Selektion, solange die Strukturen nicht entstehen, die ausgelesen werden sollen.
3.
Es liegt eine klare Diskontinuität von B nach C vor: Es bedürfte Hunderter von Zwischenstufen, um vom gelartigen Glaskörper (vitreous mass) durch Mikromutationen und Selektion zum Linsenauge von Tridacna maxima zu gelangen: Das Gel müsste schrittweise reduziert und die zelluläre Linse über viele Zwischenstufen aufgebaut werden. Hier stellt sich wohl auch schon das Synorganisationsproblem: Aufbau und Reduktion sollten bei vielen Schritten auf die Dauer aufeinander abgestimmt sein. Weiter erhebt sich die Frage nach dem Selektionswert solcher Zwischenstufen. Sollen die ersten kleinen Schritte zur Linsenbildung bereits über Tod und Leben ganzer Populationen und Organismengruppen entscheiden, wie uns das Darwin (zitiert pp. 58/59) zeigt?
Und wie soll die Selektion folgendes Problem gelöst haben: "Making the eye larger and the aperture smaller will improve spatial resolution until the aperture becomes so small that the diffraction limit is reached. After that, smaller apertures will make the image worse, not better"? (Land 1981, pp. 509/510)
Selbst Hoimar von Ditfurth, der in seiner Darstellung des Themas bis dahin treu darwinschen Gedankenlinien gefolgt war, und der glaubt, dass aus einem "ganz äußerlichen, eigentlich nur mechanischen Grund" (nämlich der Verstopfung des Lochauges durch Fremdkörper - "Nautilus könnte ein Lied davon singen") jetzt von der Selektion Mutanten begünstigt wurden, "bei denen das Loch des Lochauges vom Rand her durch eine Fortsetzung der Körperhaut verschlossen wurde", fährt 1976, p. 123, fort:
Natürlich nur dann, wenn das Häutchen sehr dünn und pigmentarm war, so daß es immer noch Licht hindurch ließ.
Trotzdem muß an diesem Punkt die Funktion des Lochauges vorübergehend beeinträchtigt gewesen sein. Daß ein solcher häutiger Verschluß in seinen Anfangsstadien den Camera obscura-Effekt nicht herabgemindert haben sollte, ist nicht vorstellbar. Die Verschlechterung der Funktion wurde offensichtlich aber wettgemacht durch den Schutz des bis dahin so anfälligen Auges. Die Vorteile für die Besitzer "geschlossener Lochaugen" müssen sogar von Anfang an überwogen haben, sonst wäre die Entwicklung an dieser Stelle unweigerlich abgebrochen.
Wir beobachten an dieser Stelle wieder die übliche evolutionistische Methodik: Die Evolution wird als Tatsache vorausgesetzt und daraus folgt, dass die Vorteile...von Anfang überwogen haben müssen, denn sonst hätte es ja keine weitere Evolution gegeben!
Die Frage, warum unter diesen Voraussetzungen Nautilus nach Hunderten von Millionen von Jahren ununterbrochener Augenvervollkommnung durch Mutation und Selektion nicht den Sprung zum "geschlossenen Lochauge" mit nachfolgender Linsenbildung geschafft hat, erklärt er nicht (vgl. auch Dawkins, unten).
Nach H. v. Ditfurth haben es überdies Mutation und Selektion in all den Jahrmillionen nicht einmal geschafft, das Lochkameraauge von Nautilus zum Schutz mit einem gelartigen Glaskörper zu füllen, wie wir das bei den Muscheln Arca und Lima gesehen haben (vgl. p. 90).*
Könnte es aber nicht auch so sein, dass die von Ditfurth und anderen zitierten Ursachen für die Bildung der verschiedenen Augentypen gar nicht ausreichen? Auf diesen Gedanken kommt der Autor nicht. Der von ihm postulierte Selektionsvorteil für den Schritt vom Lochkameraauge zum geschlossenen Auge kann jedoch für den obigen Fall der Muscheln nicht zitiert werden: Ihr Lochkameraauge war durch den gelartigen Glaskörper geschützt.
Aber auch Salvini-Plawen und Mayr bemerken Seite 249, dass sie bei der Frage nach dem genauen Selektionsdruck Schwierigkeiten bei den verschiedenen Augenformen haben.
*V.C. Barber berichtet 1987, p. 226: "Earlier descriptions of the eye of Nautilus suggested that the retina is in direct contact with the seawater. I found that the interior of the eye was filled with a jellylike substance that could have been produced by the mucos cells that occur near the edge of the retina." Der Autor fährt dann jedoch fort: "This may not be the case, since an exchange of fluid between the ocean and the interior of the eye may occur." Siehe auch Huntz 1987, Carlson 1987 (alle Beiträge herausgegeben von Saunders und Landman 1987).
4.
Die postulierten Schritte, die durch Mikromutationen und Selektion mehrfach zum selben Ziel geführt haben sollen, müssten durch diese Faktoren auch prinzipiell reproduzierbar sein.
5.
Eine weitere Diskontinuität liegt von C nach D vor: Die Lage der inversen Retinazellen kann man schlecht mit einem Schritt von der eversen Form ableiten. Bei einer Umwandlung durch Mikromutationsschritte bedarf es auch hier Hunderter Zwischenstufen. Weiter erhebt sich an dieser Stelle wieder die Frage nach dem Selektionswert solcher Zwischenstufen: Wie wir oben gesehen haben, soll ja nach Bakken und vielen anderen Neodarwinisten gerade die inverse Lage der Retina ein Musterbeispiel miserabel zusammengestümperter ("badly bungled") Strukturen bei den Organismen sein, die aus diesem Grunde nicht für die Weisheit und Schöpfung Gottes eingesetzt werden können. Und diese Fehlkonstruktionen von inversen Retinarezeptoren sollen in Form vieler kleiner Schritte jeweils klare Selektionsvorteile gegenüber der besseren eversen Retinalage gehabt haben? Wenn die Lichtrezeptoren jedoch in ihrer eversen Lage optimal funktioniert haben - welche Vorteile hatte dann diese ganze (Fehl-)Entwicklung?
6.
Um in den Rang naturwissenschaftlicher Tatsachen gelangen zu können, müsste auch dieser postulierte Umwandlungsprozess über viele Zwischenstufen durch Mikromutationen, Rekombination und Selektion prinzipiell reproduzierbar sein, zumal sich solche Vorgänge schon vielmals in der Natur abgespielt haben sollen.
7.
Da der globale Vervollkommnungsprozess der Augenentwicklung bei diesen und anderen Formen schon seit Hunderten von Jahrmillionen abgelaufen sein soll, warum haben dann A und B immer noch keine Linse entwickelt und warum ist bei C die Lage der Retina immer noch evers? Warum haben überdies D und E noch keine Iris, Fokussiereinrichtungen, das Pekten des Adlerauges etc. entwickelt, d.h. warum ist die gesamte Feindifferenzierung noch so weit von der eines Falken- oder Adlerauges entfernt?
Dawkins stellt als Neodarwinist eine ähnliche Frage für das Auge von Nautilus (1986, p. 85/86):
...Nautilus is a bit of a puzzle in its own right. Why, in all the hundreds of millions of years since its ancestors first evolved a pinhole eye, did it never discover the principle of the lens? The advantage of a lens is that it allows the image to be both sharp and bright. What is worrying about Nautilus is that the quality of its retina suggests that it would really benefit, greatly and immediately, from a lens. It is like a hi-fi system with an excellent amplifier fed by a gramophone with a blunt needle. The system is crying out for a particular simple change. In genetic hyperspace, Nautilus appears to be sitting tight next door to an obvious and immediate improvement, yet it doesn't take the small step necessary. Why not? Michael Land of Sussex University, our foremost authority on invertebrate eyes, is worried, and so am I. Is it that the necessary mutations cannot arise, given the way Nautilus embryos develop? I don't want to believe it, but I don't have a better explanation.
Es werden die erstaunlichsten Möglichkeiten in Betracht gezogen, nur nicht, dass Mutation und Selektion für den Ursprung neuer synorganisierter Systeme unzureichend sein könnten. Wenn aber die notwendigen Mutationen in diesem Beispiel in Hunderten von Jahrmillionen de facto nicht aufgetreten sind, dann zeigt das eben sehr deutlich, dass für eine Linsenbildung mehr notwendig ist als nur ein "particular simple change". Es handelt sich statt dessen um einen genetisch, biochemisch, ontogenetisch und morphologisch hochkomplexen Strukturbildungsprozess, für dessen raum-zeitliche Integration in ein bestehendes System Dutzende (vielleicht sogar Hunderte?) von genauestens aufeinander abgestimmten Mutationsschritten erforderlich wäre. Die Synorganisationsfrage ist dabei für das Gesamtsystem noch gar nicht gestellt. Die neodarwinistischen Simplifikationen gehen an den naturwissenschaftlichen Realitäten und Problemen vorbei.
Zurück zu unserem Ausgangspunkt p. 90:
8.
Bei D und E ist die räumliche Struktur und Anordnung von Linse und Netzhaut neu aufeinander abgestimmt. Hier stellt sich wieder das Synorganisationsproblem. Wiederholte Entstehung solcher Synorganisationen durch Mikromutationen und Selektion? Warum ist man dann nicht in der Lage, die postulierten Prozesse zu reproduzieren?
9.
Salvini-Plawen und Mayr schreiben weiter auf p. 238:
The latter eyes (those of Tridacna) have been investigated ultrastructurally by Kawaguti and Mabuchi (1969) and show some rhabdomeric receptorial cells and a predominance of ciliary photoreceptors. In the siphonal eyes of Tridacna as well as of Cardium (both Cardiacea), the optic nerve divides and innervates also a so-called ciliary sense organ (ciliated receptor) close to the eye. (Vgl. Abb. p. 90)
Zum letzteren Punkt: Wie soll man sich die Teilung des Sehnervs und die Bildung des ziliaren Sinnesorgans in vielen kleinen Mutationsschritten, die nur "slight or even invisible effects on the phenotype" (Mayr 1970, p. 169, ähnlich 1979, pp. 40/41) haben sollen, vorstellen und welche entscheidenden Selektionswerte sollen alle diese kleinen Schritte gehabt haben? (Dokumentation der Behauptungen im Detail siehe Lönnig 1988.)
Auch dieser Entwicklungsschritt sollte reproduzierbar sein.
10.
Fortsetzung des Abschnittes von p. 238:
The respective homology is obvious and clarifies the origin of the inversion in the Cardium eye.
Das Letztere ist falsch: Das Cardium-Auge hat nach der Abbildung auf p. 90 noch das ziliare Sinnesorgan und die Retinalage ist bereits invers.
11.
Fortsetzung des Abschnittes von p. 238:
The ultrastructure of the retinal cells shows, as in Tridacna, photoreceptors of the ciliary type with (9 x 2) + 0 axonemes of the 200 - 300 cilia a cell (Barber and Wright, 1969); the ciliary sense organ bears cilia with (9 x 2) + 2 microtubules without folding of the ciliar membrane
In comparing the respective organs of Cardiacea (Tridacna, Cardium) and of Pectenacea (Lima, Spondylus, Pecten) the almost complete identity of the eye in Pecten and in Cardium is obvious (Küpfer, 1916) - the ciliary sense organ in Cardiacea, however, has become in the eye of Pecten the distal retina by inversion. (Siehe Abb. p. 90 D und E)
Der Stil des letzten Satzes erinnert mich an Aussagen wie: "Sie waren beide fast vollkommen gleich groß, nur der eine war wesentlich größer als der andere".
Zu den Unterschieden nun wieder die bekannten Fragen:
a)
Wie ist das ziliare Sinnesorgan (das bei den Pectenacea überhaupt nicht auftaucht!) bei Pecten maximus durch viele hundert Mikromutationsschritte, mit geringfügigen oder sogar unsichtbaren Wirkungen auf den Phänotyp zur distalen Retina geworden? (Oder soll das mit einem Schwung geschehen sein? - Das wäre jedoch bei der komplizierten raum-zeitlichen und funktionellen synorganisierten Neuorientierung großer Teile des Gesamtsystems denkbar unwahrscheinlich!)
b)
Nachdem die Selektion nun in mühevoller Kleinarbeit unter ungeheuren Opfern (vgl. Darwin, zitiert pp. 48/49, dass alle alten Zustände des Instruments zerstört werden) das ziliare Sinnesorgan schrittweise aufgebaut und bis zum fertigen Zustand vervollkommnet und damit dann endlich den optimalen Funktions- und Selektionswert erreicht hatte, baut sie es - wieder unter ungeheuren Opfern von Millionen und Milliarden von Individuen vieler Generationen über Hunderte von Mikromutationsschritten in die distale Retina um?! Selektionstheoretisch wäre es jedoch viel günstiger gewesen, den Sehnerv einfach noch einmal zu teilen und die distale Retina unabhängig von dem ziliaren Sinnesorgan aufzubauen. Aber auch hier würde sich wieder die Frage nach den Selektionswerten der einzelnen Schritte stellen und das Synorganisationsproblem ist dabei noch gar nicht genannt.
Selbst Neodarwinisten halten ihre Evolutionsfaktoren eines göttlichen Schöpfers für unwürdig. So lesen wir bei Dobzhansky, Boesiger und Sperlich 1980, p. 136: "Wenn ein Gott den Plan der gesamten Evolution nur dazu ausgedacht hätte, um schließlich den Menschen daraus hervorgehen zu lassen, so müßte man von einer ganz erstaunlich ineffektiven Planung sprechen. Warum müssen hierfür zwei bis drei Milliarden Jahre vergeudet werden? Warum all dieses Leiden, diese Schwierigkeiten und das Entstehen und Vergehen von vielen Millionen von Arten, um zu diesem Ziel zu gelangen?"
c)
Die Frage nach dem Selektionswert der einzelnen kleinen Schritte stellt sich natürlich ebenso für die direkte Ableitung der distalen Retina vom ziliaren Sinnesorgan.
d)
Es erhebt sich wieder die Frage nach der Reproduzierbarkeit der postulierten Umwandlungs- und Vervollkommnungsprozesse.
e)
Pectens Linsenform unterscheidet sich klar von der Cardiums. Zur Ableitung durch Mikromutationsschritte bedürfte es vieler Übergangsstufen.
f)
Zwischen den Augen von Lima und Pecten (beides Pectenacea) besteht eine große Kluft (weitere Diskontinuität). Sie soll nach Salvini-Plawen und Mayr durch die Cardiacea überbrückt werden (Hilfshypothese!). Die Letzteren weisen jedoch vorherrschend (Tridacna) oder ausschließlich (Cardium) den ziliaren Rezeptortypus auf, während Lima (überhaupt?) und Pecten in der proximalen Retina den rhabdomeren Typus zeigen, aber Cardium darüber hinaus noch den ziliaren Typ der Lichtrezeptoren in der distalen Retina hat. Wenn Letzterer (E) aber der Vorläufer von C und D war, dann treten in dieser Serie wahrhaftig die Kinder vor den Eltern auf! Bei der ganzen Ableitung stört nur das bei C und D noch vorhandene ziliare Sinnesorgan, das sich ja in die inverse Retina umgewandelt haben soll.
Dass die Autoren tatsächlich diese Ableitung vornehmen, zeigen sie auch in ihrer Zusammenfassung p. 247:
...it appears that here the rhabdomere type constitutes the general condition of currently prevailing photoreceptoral organization, which in the course of evolutive improvement has been either supplemented by cells of the ciliary type or been replaced altogether by receptors of the ciliary type. Tridacna, with unquestionably rather primitive everse eyes, still shows a mixture of rhabdomeric and (prevailing) ciliary type...The inverse eyes of Cardium are exclusively ciliary. This conclusion is supported by the structure of photoreceptors in Pecten, the proximal, original retina of which shows rhabdomeric pattern (Barber et al., 1967), whereas the new distal retina possesses ciliary structure and is the real image-receptoral layer* (Land, 1968, p. 81).
Die Serie müsste demgemäß wie folgt aussehen: A, B, E, C, D - womit die Unterschiede noch deutlicher hervortreten.
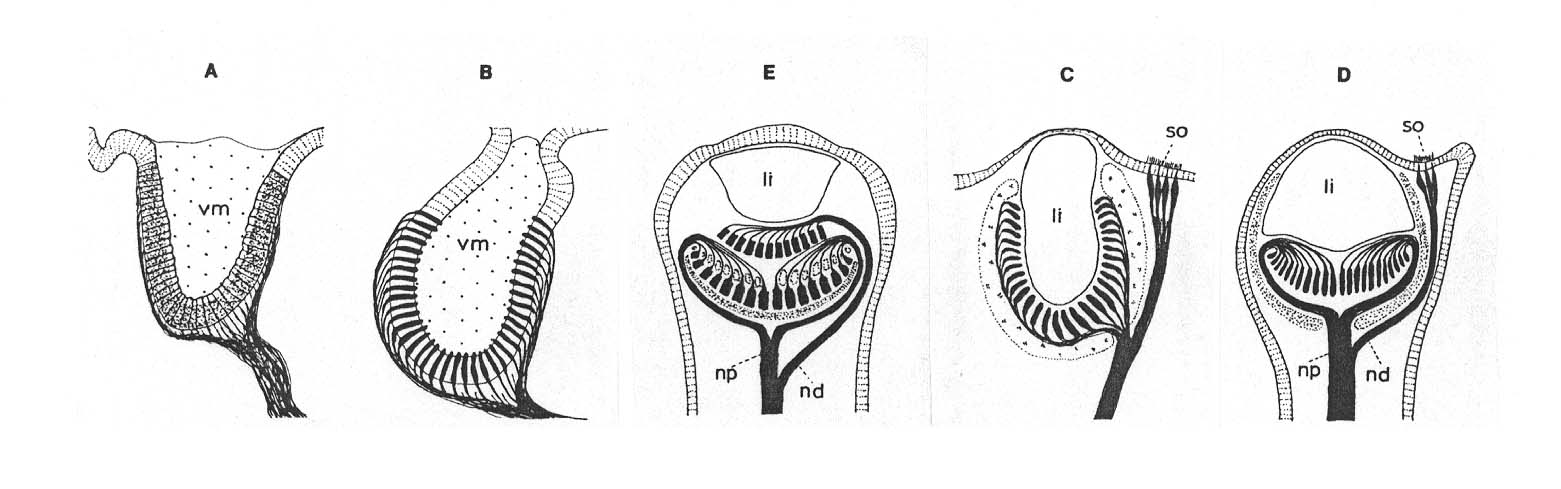
Aus Salvini-Plawen und Mayr. Reihenfolge nach biologischen Fakten verändert!
*"Hartline (1938) had shown that receptors in this layer responded to the offset of light, whereas receptors in the proximal retina - which is adjacent to the argentea and does not receive a focussed image - respond to the onset" (Land 1981, p. 536; dort weitere Details). Beide Retinae haben Funktionen.
12.
Zu Pectens distaler Retina bemerken die Autoren außerdem in der Fortsetzung des Abschnittes von p. 238:
This distal retina characteristically shows (besides some retained microvilli) the ciliary type of photoreceptoral structure in sacs ((9 x 2) + 0 axonemes...), whereas the original proximal retina is differentiated into the rhabdomeric type (Eakin 1968). Thus, there are two recent well-known sets of photoreceptors at the mantle edge of bivalves, the Cardiacea line (Tridacna to Cardium) and the Pectinacea line (Lima to Pecten) both of which might have a joint route at such a simple level as still represented in the Arcacea (see also Küpfer, 1916). The morphological gap existing within the Pectinacea line (between Limidae and Pectenidae) is almost completely bridged by the Cardiacea line (Fig.) and the whole sequence supports the important conclusion that sensory cells of one type can be replaced or supplemented by cells of another type (p. 247).
Schon bei den Ausführungen Salvini-Plawens und Mayrs, zitiert p. 93 unten, wird sich der eine oder andere Leser gefragt haben, warum man überhaupt Photorezeptoren des Ziliartyps ("9 x 2 + 0 axonemes of the 200 - 300 cilia a cell") feststellen kann, wenn diese gar nichts mit dem ziliaren Sinnesorgan zu tun haben.
Bei solchen strukturellen Ähnlichkeiten sprechen Salvini-Plawen und Mayr auch häufig von Rudimenten, um damit den Evolutions- und Abstammungsgedanken zu verstärken. Rudiment meint dabei in der Regel das funktionslose, nur historisch zu erklärende Überbleibsel der phylogenetischen Entwicklung. Das grundsätzliche Problem ist dabei jedoch, wie nach Hunderten von Jahrmillionen ununterbrochener Verbesserungsarbeit durch Mutation und Selektion überhaupt noch funktionslose Strukturen vorhanden sein können. Die Evolutionsfaktoren sollen ja auch die kleinsten Details der Entwicklung nicht unberücksichtigt lassen - ist doch die natürliche Selektion "täglich und stündlich dabei, überall in der Welt die geringsten Veränderungen aufzuspüren und sie zu verwerfen, sobald sie schlecht sind, zu erhalten und zu vermehren, sobald sie gut sind". - Darwin 1872/1967, pp. 125/126.
Kein Geringerer als Autrum schreibt 1979, p. 9:
We are left with the unsolved Problem of why so many different photoreceptors have ciliumlike structures and what their function is. The inference that they are only a "relict", as Vinnikov (1974) maintains, is unsatisfactory indeed."
Es wurde schon hervorgehoben, dass das ziliare Sinnesorgan und der ziliare Photorezeptortyp zusammen bei bestimmten Muscheln auftreten, so dass eine Ableitung des einen vom anderen nicht geboten erscheint. Verstärkt wird diese Auffassung noch durch die Schwierigkeiten und Widersprüche, die einer Ableitung durch Mikromutationen und Selektion entgegenstehen (s.o.).
Sieht man sich weiter in der unbelebten und lebenden Schöpfung gründlich um, dann stellt man fest, dass bestimmte gleichartige hochqualifizierte und erprobte Struktur- und Bauelemente immer wieder von neuem an den verschiedensten Stellen für bestimmte verwandte Funktionen eingesetzt werden. (Im Bereich der Technik finden wir Ähnliches etwa bei den Mikrochips in Schreibmaschinen, Videokameras, Computern etc.). Man kann vielleicht von einer ideellen Ableitung der Strukturen zur Bewältigung ähnlicher Aufgaben sprechen.
Richtig müsste der ganze Absatz Salvini-Plawens und Mayrs von p. 238 etwa wie folgt lauten:
The sequence from epidermal photoreceptors (Arca) to almost closed everted eyes (Lima/Pectenacea)* and to the closed eyes with a multi-cellular lens (Tridacna; Stasek, 1966) offers a number of heavy and inextricable problems for an evolutionary interpretation by means of mutations and selection: The evolutionary process is highly improbable; none of the postulated transformations is reproducible. The eyes of Tridacna have been investigated ultrastructurally by Kawaguty and Mabuchi (1969) and show a predominance of ciliary photoreceptors and some rhabdomeric receptoral cells. In the siphonal eyes of Tridacna as well as of Cardium (both Cardiacea)**, the optic nerve divides and innervates also a so-called ciliary sense organ (ciliated receptor) close to the eye (Abb. p. 90). The respective homology of this sense organ is obvious, but has nothing to do with the origin of the inversion in Cardium, as both - the inverted retina as well as the ciliary sense organ, exist side by side in the same organism. The ultrastructure of the retinal cells of Cardium shows, as in Tridacna, photoreceptors of the ciliary type with (9 x 2) + 0 axonemes of the 200 - 300 cilia a cell (Barber and Wright, 1969); the ciliary sense organ bears cilia (9 x 2) + 2 microtubules without folding of the ciliar membrane. In comparing the respective organs or Cardiacea (Tridacna, Cardium) and of Pectenacea (Lima, Spondylus, Pecten) the strong difference between the eyes of Cardium and Pecten is obvious: instead of the ciliary sense organ in Cardiacea, Pecten has a distal inverted retina. As the improbability of a transformation by many hundreds of micromutations with no selection value and non-functioning intermediate links in a slow evolutionary process of millions of years is evident and as a macromutation is equally absurd, an accidental derivation of the inverted retina from the ciliary sense organ is hardly possible. The connection in creation is, however, obvious: The Creator programmed the optic nerve to divide and grow into the ciliated sense organ in the Cardiacea and to divide and develop into the distal inverted retina in Pecten. For its special functions this distal retina shows (besides some microvilli), the ciliary type of photoreceptoral structure in sacs ((9 x 2) + 0 axonemes), whereas the proximal retina is differentiated into the rhabdomeric type (Eakin, 1968). Thus, there are two complex and well-known sets of photoreceptors at the mantle edge of bivalves, in the Cardiacea (as Tridacna and Cardium) and the Pectinacea (as Lima and Pecten), both of which show huge anatomical gaps between their members (instead of an evolutionary continuum) and none of which might have a phylogenetic joint route at a less complex level as is represented in the Arcacea (in contrast to Küpfer, 1916). The morphological gap existing within the Pectinacea (between Limidae and Pectinidae) is absolutely not bridged by the Cardiacea forms [Abb. p. 90 und p. 97] and the different other anatomical and functional gaps support the important conclusion that sensory cells of one type cannot systematically be replaced or supplemented by cells of another type by micromutations and selection, but only by intelligent direction.
*Der Einfachheit halber habe ich die veraltete Systematik von Salvini-Plawen und Mayr im Folgenden beibehalten.
**Der Gedanke kommt mir an solchen Stellen, dass die Autoren diese Systematik vor allem deshalb gebrauchen, um dem Leser ihre Abstammungshypothesen näher zu bringen. A. H. Müller benutzt die neuen Familienabgrenzungen hingegen bereits 1956 in seinem Lehrbuch der Paläozoologie (l. Aufl.) und Salvini-Plawen gebraucht in seinem Beitrag zu Grzimeks Tierleben 1975/1977 ebenfalls die neue Systematik. Aber die alte Systematik rettet die Sache auch nicht mehr.
In ähnlicher Weise könnte man den gesamten Text Salvini-Plawens und Mayrs korrigieren.
Nach Diskussion der obigen morphologischen Serie dürfen wir zunächst einmal unsere bisherige Schlussfolgerung weiter begründet aufrechterhalten:
The modern evidence showing that Darwin was wrong is overwhelming.
Die eben behandelte morphologische Serie bei den Bivalvia (Muscheln) ist als Evolutionsbeweis so schwach, dass selbst Bakken diese Serie weglässt und nur von "two such graded series of eyes in living prosobranch gastropods and polychaete worms" spricht. Diesen beiden Serien wollen wir uns als nächstes zuwenden.