WEITERE DISKUSSION
VORBEMERKUNGEN: PALÄONTOLOGIE UND MAKROEVOLUTION
W.-E. L.
Was hier manchem Leser auf den ersten Blick vielleicht als überzeugende Beschreibung der Mikroevolution erscheint (mit Extrapolationsmöglichkeiten für makroevolutionistische Fragestellungen), erweist sich bei genauerer Betrachtung in allen wesentlichen Punkten als problematisch und ist als Ansatz für die postulierte Makroevolution unzutreffend.
Bevor ich Prof. K.s Ausführungen Punkt für Punkt diskutiere, möchte ich die neodarwinistische Problematik anhand des Kambriumproblems verdeutlichen. Ich zitiere im folgenden die Ausführungen der evolutionistischen Autoren Robert Augros und George Stanciu (1988, pp. 219, 221/22) mit der Bitte an den (mit dem Problem vielleicht noch nicht völlig vertrauten) Leser, zunächst einmal diesen für unsere Fragestellung aufschlussreichen Text gründlich durchzustudieren:
Der Fossiliennachweis über die gesamte geologische Geschichte zeigt eine langfristige Abnahme der Zahl der Stämme und anderer höherer Kategorien, obwohl die Zahl der verschiedenen Arten insgesamt zunimmt. Das Ergebnis ist eine intensivere Diversifizierung in immer kleineren Grenzen. Gould nennt als Beispiel die Meerestiere: "In unseren modernen Meeren dominieren nur noch wenige Gruppen - vor allem Muscheln, Schnecken, Krabben, Fische und Seeigel-, aber jede umfasst mehr Arten, als irgendein Stamm im Paläozoikum jemals erreichte (vielleicht mit Ausnahme der Trilobiten im Ordovizium und der Seelilien im Karbon). Paläozoische Meere enthielten vielleicht nur halb so viele Arten wie die heutigen Ozeane, doch besaßen diese eine große Vielfalt grundlegend verschiedener Baupläne. Diese stetige Abnahme der Mannigfaltigkeit von Bauplänen - bei gleichzeitiger starker Zunahme der Artenzahl - stellt wohl den außergewöhnlichsten Trend des Fossiliennachweises dar."
Der Zoologe James Brough von der Universität von Wales betrachtet die "Evolution als einen Prozess, der sich stetig verlangsamt". Er weist darauf hin, dass alle heute noch lebenden Tierstämme seit dem Kambrium existieren, also seit 500 Millionen Jahren. Seither sind keine neuen Tierstämme entstanden, doch einige der frühen sind inzwischen ausgestorben. In ähnlicher Weise treten seit dem unteren Paläozoikum, seit ungefähr 400 Millionen Jahren, keine neuen Klassen innerhalb dieser Tierstämme auf. Alle heutigen Klassen kamen zusammen mit anderen vor, die nicht bis zur Gegenwart überlebt haben. Die Bildung neuer Ordnungen hörte gegen Ende des Mesozoikums auf, vor ungefähr 60 Millionen Jahren. Brough konstatiert auch eine Abnahme im Auftreten neuer Familien und kommentiert den Trend folgendermaßen: "Die Evolution war in einer Reihe immer stärker eingeschränkter Gebiete wirksam, und die Auswirkungen im großen Maßstab gingen immer mehr zurück...In Zukunft wird die Evolution wohl auf immer kleineren Gebieten wirksam werden, bis sie vollig aufhört." Dieser Trend deutet darauf hin, dass die Evolution keine ewige, unbegrenzte Kraft darstellt, sondern selbst Beschränkungen unterliegt und sich in einem langen, aber begrenzten Zeitraum abspielt. Der zweite Hauptsatz der Thermodynamik sagt, dass das Universum als Gesamtes seinem Ende entgegengeht. Der Fossliennachweis behauptet dasselbe für den Evolutionsprozess.
Diese "Verschiebung von wenigen Arten in vielen Gruppen zu vielen Arten in wenigen Gruppen" widerspricht rundweg dem Darwinschen Gradualismus. Denn wenn die Evolution so voranschritte, dass die Arten kleine Veränderungen summieren, dann müssten über lange Zeiträume hinweg mit zunehmender Häufigkeit neue Ordnungen, Klassen und Tierstämme auftreten. Aber gerade das Gegenteil geschieht bei den Fossilien. Darwins Modell ist verkehrt. (Hervorhebungen im Schriftbild von mir.)
Der (ebenfalls evolutionistische*) Paläontologe S.J. Gould hat zu dieser Frage 1989 eine umfassende Monographie vorgelegt (347 pp.), in der er die nach Augros und Stanciu oben zitierten Aussagen noch einmal in allen Details bestätigt. Tatsächlich ist das Kambriumproblem für den Neodarwinismus durch die Identifizierung zahlreicher weiterer kambrischer Tierstämme in den letzten Jahren noch brisanter geworden! Alle Formen treten sprunghaft auf und nach Auffassung führender Evolutionsforscher fehlen damit von vornherein fünf Sechstel (A. Watson 1987) bis sieben Achtel (Schopf 1987) des für eine kontinuierliche Evolution erhofften Belegmaterials. (Die Ediacara-Fauna ist dabei zu einem Sonderproblem geworden, statt das Problem zu lösen.) Den generellen Kontrast zwischen neodarwinistischen Prognosen und paläontologischen Realitäten hat Schindewolf 1965, pp. 85/86 treffend auf folgenden Nenner gebracht (ähnlich Valentine und Jablonsky 1987, Bottjer 1987, Meyen 1988, Willmer 1990, Bengtson 1990, Devillers, Chaline und Laurin 1990 u.v.a.):
"Nach darwinistischer Vorstellung sollen geringfügige Rassenunterschiede sich allmählich zu Artmerkmalen verstärken und diese dann durch Addition immer neuer kleiner Abänderungen zu Gattungs-, Familienunterschieden und so weiter werden. Die Formenmannigfaltigkeit müsste alsdann gegen Ende der einzelnen Stämme zunehmen; dort wäre die größte Fülle von Ordnungen, Familien und Gattungen, das heißt von Unterschieden höheren Grades zu erwarten. Das Gegenteil ist der Fall.
Ein neuer Bauplan von dem systematischen Range etwa einer Klasse oder Ordnung erscheint gewöhnlich völlig unvermittelt auf der Bildfläche, ohne lange Reihen von Bindegliedern, die uns eine allmähliche Herausgestaltung aus einer anderen, seine Wurzel bildende Ordnung vor Augen führen würden."
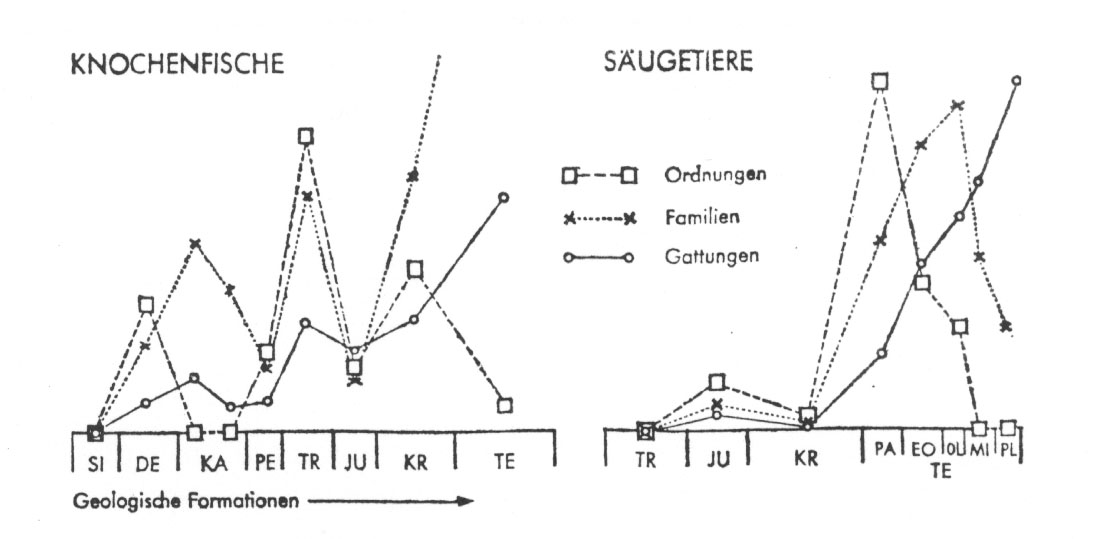
Simpson und Schindewolf haben außerdem gezeigt, dass die Maxima des Auftretens höherer systematischer Kategorien denen der niederen regelmäßig vorausgehen.
Kurven zeitlicher
Maxima in der Hervorbringung neuer Ordnungen, Familien und Gattungen
innerhalb der Klassen der Knochenfische (Osteichtyes. links) und der
Säugetiere (Mammalia, rechts). (Nach Simpson.) - Die Abszisse
verzeichnet die Folge der geologischen Formationen vom Silur bzw. von der Trias bis zum Tertiär, letztere im Falle der Säugetiere noch unterteilt in die einzelnen Stufen. In der Ordinate kommen die relativen Zahlen der neu auftretenden Systemgruppen zum Ausdruck. Die Schemata beruhen auf genauen statistischen Auszählungen; die absoluten Zahlenwerte sind ohne Interesse. - Die Höhepunkte in der Produktion neuer Ordnungen gehen im allgemeinen sehr deutlich denen der Familien und diese wiederum denen der Gattiugen voraus. Bei wachsender morphologischer und systematischer Divergenz wäre ein umgekehrtes Bild zu erwarten: am Anfang nur wenige indifferente Ordnungen und Familien, eine ständige Zunahme ihrer Zahl gegen Ende der Stammesentfaltung, wobei eine zeitliche Folge der Zahlenmaxima von den Gattungen zu den Familien und schließlich zu den Ordnungen hin auftreten müsste.
Aus Schindewolf 1965, p. 88 (Hervorhebungen im Schriftbild von mir).
1987 und 1988 haben mehrere Paläontologen diese Befunde noch einmal bestätigt (Erwin, Valentine, Jablonsky, Bottjer; vgl. Lewin 1988, pp. 292/293: ""The fossil record suggests that the major pulses of diversification of phyla occurs before that of classes, classes before that of orders, and orders before that of families", say Erwin and his collegues. "This is not to say that higher taxa have originated before species..., but the higher taxa do not seem to have diverged through an accumulation of lower taxa. ...Jablonsky and Bottjer were able to show that the pattern is not an artifact of preservation: it is real and therefore must be saying something about evolutionary mechanisms. The most obvious message is that a simple extrapolation from one level to another is an unlikely explanation of evolutionary innovation" (Kursiv von mir.)
Die neodarwinistische Hilfshypothese von der allgemeinen "Lückenhaftigkeit der Fossilüberlieferung" wird überdies durch das (schon in der Fußnote des vorigen Kapitels erwähnte) Faktum von wenigstens 100 Millionen in den Museen der Welt katalogisierten (Makro-)Fossilien widerlegt (Kier 1981, Smithonian Institution, Washington; Raup vom Field Museum in Chicago spricht 1979 sogar von 200 Millionen. Dazu kommen die in die Milliarden gehenden Mikrofossilien). Auf weitere Details zu dieser Frage möchte ich bei der Analyse von Prof. K.s Ausführungen zurückkommen.
Die Problemlage ist damit klar umrissen: 1. Seit dem Kambrium ist eine stetige Abnahme der Mannigfaltigkeit der Baupläne und anderer höherer Kategorien festzustellen. 2. Trotz eines ungeheuer reichen Materials bei gut fossilisierbaren Formen erscheinen auch die höheren Taxa solcher Gruppen gewöhnlich völlig unvermittelt auf der Bildfläche. 3. Die höheren Kategorien (d.h. die großen morphologisch-anatomischen und physiologischen Divergenzen von Bauplänen, Klassen und Ordnungen) haben ihre zeitlichen Maxima regelmäßig vor den niederen systematischen Einheiten.
*Das trifft auch auf die meisten der folgenden zitierten Autoren zu.