A. RESISTENZ-PHÄNOMENE
Resistenz-Phänomene werden in vielen Lehrbüchern als Musterbeispiele der Evolution neuer lebenswichtiger Eigenschaften und Fähigkeiten durch Mutation und Selektion beschrieben. Verschafft man sich einen Überblick über die Primärliteratur zu dieser Frage, so wird zunächst die vielfach zu lesende Aussage bestätigt, dass die Resistenzallele in den Populationen in der Regel bereits vorhanden sind. Die Feststellung wird in der Sekundärliteratur häufig unter dem Begriff der Präadaptation geführt.
Wir möchten zunächst festhalten, dass diese Tatsache in voller Übereinstimmung mit der Regel der Rekurrenten Variation ist. Bei den oben zitierten Mutationsraten muss bei größeren Populationen praktisch zu jeder Zeit eine enorme Zahl mutierter Gene vorliegen, und das Mutantenspektrum muss sich über einen großen Teil von Individuen erstrecken, deren Fitness weit unter der des optimal angepassten und funktionierenden Genotyps liegt.
Zweitens ist hervorzuheben, dass das Resistenz-Phänomen eine Palette verschiedener genetischer Ursachen umfasst, die wir nach summarischer Aufführung näher betrachten möchten. Zu diesen Ursachen gehören:
1. Punktmutationen
a) (meist) rezessiv (Strukturabbau)
b) (seltener) dominant (z.T. Rückmutationen: Strukturwiedergewinn)
2. Rekombination (komplementäre Genwirkungen)
a) vermehrte oder
b) verminderte Bildung bestimmter für die Resistenz relevanter Verbindungen
3. Genamplifikation
4. Ploidiegrad
5. Transposons und IS Elemente
6. Plasmide (R-Faktoren)
7. Selektion von Repair-Systemen
8. Cytoplasmatisch bedingte Resistenz (Chloroplasten und Mitochondrien)
9. Resistenz durch Transposon-vermittelte Promotorsequenzen für die Expression stiller Gene (für Eukaryonten noch hypothetisch)
Zunächst zu den Punktmutationen, deren Anteil an neu auftretenden Resistenz-Phänomenen bei Eukaryonten weit über 80 Prozent der Fälle betragen dürfte: Viele Leser werden sicherlich erstaunt sein zu erfahren, dass es sich auch hier in der großen Mehrheit um Abbauprozesse handelt. Diesen Punkt möchte ich mit Beispielen aus der Primärliteratur im folgenden belegen.
Ashburner et al. berichten 1982, dass sie bei Drosophila melanogaster 28 Röntgenstrahlen-induzierte und 2 spontane Adh (Alkoholdehydrogenase) Null-Mutanten aufgrund der Resistenz gegen die toxischen Effekte von 1-Penten-3 ol [die Mutanten sind also resistent gegen 1-Penten-3 ol, weil sie die Alkohole nicht mehr umsetzen können] selektierten (bei den 28 Röntgenstrahlen-induzierten Mutanten handelte es sich in 20 Fällen um Deletionen des Adh Locus und wenigstens ein benachbartes Gen, - was auch auf eine der beiden Spontanmutanten zutraf).
Braaksma und Feenstra fassen 1982, p. 83, ihre Studie zur Isolation und Charakterisierung von Nitrat-Reductase Mangelmutanten bei Arabidopsis thaliana wie folgt zusammen (Auszug):
Chlorate resistent mutants of Arabidopsis thaliana were isolated, of which 10 exibited a lowered nitrate reductase activity and 51 were chlorate-resistent because of an impaired uptake of chlorate.
Buxton und Radford berichten 1982, p. 128, über Fluoropyrimidine-resistente Mutanten von Neurospora crassa:
Mutants defective in these systems [pyrimidine metabolism: uptake and salvage] have been selected by their inability to utilize exogenously supplied pyrimidines to supplement growth of pyrimidine auxotrophs (Williams and Mitchell 1969).
(p. 191:) Mutations in at least six loci render N. crassa resistent to fluoropyrimidines. Although some of these loci are linked, they are not tightly linked to each other or to other known loci involved in pyrimidine metabolism. A mutant in uc-2...has previously been shown to lack thymidine, 2-Oxoglutarat Dioxygenase...and is FUdR-resistant and FUR-and FU-sensitive. Mutants in uc-4...are FU-resistant, FUR- and FUdR-sensitive, and lack uracil phosphoribosyl transferase...The mutant uc-5...lacks uracil uptake...A mutation in fdu-2...is resistant to FU, FUR, and FUdr and hence may be affected in regulation of the de novo biosynthesis of pyrimidines.
In ihrer Einleitung weisen die beiden Autoren darauf hin, dass die Resistenz gegen toxische Pyrimidin-Analoga durch Mutationen in mehreren Systemen hervorgerufen werden kann: Blockade der Aufnahme, Überproduktion des normalen Stoffwechselweges oder verstärktem Abbau des Analogons. In der obigen Arbeit sprechen sie nur von Mutanten, die nicht in der Lage waren, exogen zugeführtes Pyrimidin zu verwerten.
Johnston und Coddington stellen in der Zusammenfassung ihrer Arbeit MULTIPLE DRUG RESISTANCE IN THE FISSION YEAST Schizosaccharomyces pombe: CORRELATION BETWEEN DRUG AND AMINO ACID UPTAKE AND MEMBRANE ATPase ACTIVITIES (1983, p. 299) u.a. folgendes fest:
Cyh 3 and cyh 4 multiple drug resistant strains of Schizosaccharomyces pombe, show a much reduced uptake of trichodermin, chloramphenicol, cycolheximide, L-lysine, glycine, L-threonine, L-glutamine, L-arginine and L-glutamic acid when compared to wild type. The plasma membrane and mitochondrial ATPase activities are also greatly reduced. Since the uptake of such compounds is likely to be driven by a proton electrochemical gradient set up by the membrane ATPase it is suggested that the primary effect of these mutations is at the level of the membrane ATPase.
Another drug resistent strain, cyh 1,...resistant only to high levels of cycloheximide...shows an increased uptake of trichoder-min, L-lysin, glycine, L-threonine, L-glutamine when compared to the wild type.
Im letzteren Falle scheint die erhöhte Aufnahme verschiedener Stoffe die Cycloheximidverarbeitung zu begünstigen.
Kraus et al. 1979, p. 49:
Mutants of bacteria resistant to inhibition by analogues of metabolic end products have often been shown to be abnormal in regulation of the biosynthesis of the normal endproduct (Umberger, 1971).
Lengeler schreibt in seiner Arbeit CHARACTERISATION OF MUTANTS OF Escherichia coli K 12, SELECTED BY RESISTANCE TO STREPTOZOTOCIN 1980, p. 50 und p. 49 (Summary):
Antibiotics usually enter bacterial cells by means of preexisting carbohydrate-, aminoacid-, or ion-specific transport systems, whose natural substrates they mimic (Brown, 1977). Consequently, mutants deficient in one of these transport systems are resistant to an antibiotic entering through this system.
From cultures of sensitive bacteria, treated with the antibiotic streptozotocin, two classes of resistant mutants can be isolated:
1) mutants, resistant under all conditions tested to even the highest doses of the antibiotic. These are either pleiotropic-defective pts-mutants, or more frequently, mutants lacking a transport system (enzyme IINag-complex of the PEP-dependend phosphotransferase system) encoded by the gene nagE. This gene is inducible by N-acetyl-glucosamine and seems to be part of the nag operon. The transportsystem in question is responsible for the uptake of N-acetyl-glucosamine, of D-glucosamine and of streptozotocin;
2) conditional resistant mutants which are unable to energize or to synthesize the streptozotocin transport system under certain growth conditions but do have the transport activity under other conditions. These include a) mutants auxotrophic for amino acids, vitamins, or nucleotides, b) mutants negative or sensitive to carbohydrates in the medium, and c) mutants with defects in energy metabolism such as PEP synthesis.
Lewis et al. (THE GENETICS OF LEVAMISOLE RESISTANCE IN THE NEMATODE Caenorhabditis elegans) 1980:
(p. 909:) Levamisole-poisened adults crudely resemble levamisole-resistant unc mutants in motor behavior, which we interpret to mean that levamisole as an agonist interferes with the same function that is ablated to confer resistance.
Obwohl die Autoren darauf hinweisen, dass die resistenten Mutanten gute Vitalität aufweisen, werden weiter folgende Punkte hervorgehoben:
(pp. 915/916:) Apart from their pharmacological phenotype, the most obvious thing wrong with the resistant unc mutants is a discrete, rather mild defect in their motor behavior as adults. ...generally grow slightly more slowly than the wild type...have somewhat fewer progeny...Homozygous unc males... attempt copulation with less vigor and agility than wild-type males. Flexure of the male tail is more limited.
...The mild reduction in chemosensory response and male sexual behavior is consistent with loss of a noncritical component of these behaviors.
(p. 918:) Resistant unc mutants are generally more sensitive to hypo-osmotic shock than wild type.
(pp. 918/919:) Uncoordinated behavior represents loss of endogenous function, while levamisole resistance is failure to tespond to a synthetic exogenous signal.
Im Falle von RADIORESITANT DNA SYNTHESIS: AN INTRINSIC FEATURE OF ATAXIA TELANGIECTASIA (einer schweren, genetisch bedingten Muskelfunktionsstörung beim Menschen) scheint die Resistenz mit der erhöhten Replikationsrate zusammenzuhängen. Painter stellte 1981, p. 183 fest:
The radioresistance of DNA synthesis in AT cells is primarily due to a reduced inhibition of replicon initiation compared to that occurring in normal cells, but DNA chain elongation is also more radioresistant in AT cells.
Erhöhung ist hier "reduzierte Inhibition".
Roberts und Morse fassen 1980, p. 129 die Hauptpunkte ihrer Arbeit GALACTOKINASE-DEFICIENT MUTANTS OF Tetrahymena thermophila: SELECTION AND CHARACTERIZATION wie folgt zusammen:
The resistant phenotype of the mutants is apparently due to the galactokinase deficiency, which prevents the accumulation of toxic phosphorylated metabolites of 2-deoxygalactose. Genetic analysis reveal that the 2-deoxygalactose resistance alleles segregate as single Mendelian loci. The galactokinase deficient strains described here represent the first mutants in this organism for which the biochemical basis of the mutant phenotype is known.
Die Resistenz ist an rezessive Allele gebunden.
Sedgwick und Robins berichten 1980 über Resistenz gegen alkylierende Agenzien bei Escherichia coli (p. 85):
As MNNG has to be converted by thiols to the proximal mutagen methylnitrosamine, these strains were resistant simply because their low thiol levels made them less readily methylated by MNNG.
Silber und Becker kommen in ihrer Arbeit HYPOXANTHINE-GUANINE-PHOSPHORIBOSYL-TRANSFERASE (HGPRT) ACTIVITY IN THE VESTIGIAL MUTANT OF Drosophila melanogaster: EFFECT OF INHIBITORS OF THE PURINE PATHWAY zu dem Ergebnis, dass die (schwache) Resistenz der Mutanten gegen Azaserin und 6-Mercaptopurin und die bei mehreren vestigial-Mutanten ausgeprägte Resistenz gegen Aminopterin ein Resultat des gestörten Nukleotid-Metabolismus ist (1981, pp. 217 - 220).
Zahlreiche weitere Beispiele für Resistenzerscheinungen durch Strukturabbau, Störung des Metabolismus etc. sind bekannt (z.B. Wild und Klopotowski 1981, Gonzáles et al. 1981, Kraus et al. 1979, wahrscheinlich auch Maliga 1981, Nöthel 1981, Schwarz et al. 1981 u.v.a.).
Dominante Resistenzerscheinungen hingegen dürften zu einem erheblichen Teil Rückmutationen, Rekombination und möglicherweise Transposon-vermittelte Expression Stiller Gene als Ursache haben. So kann ich aus meiner Arbeit mit Antirrhinum darauf aufmerksam machen, dass mehrere Wildlinien und -arten resistent gegen den Antirrhinum-Rost (Puccinia Antirrhini) sind (Antirrhinum majus L. ssp. tortuosum, A. barrelieri, A. graniticum, A. siculum). Unsere Kulturformen sind zum größten Teil nicht (mehr) resistent. Hin und wieder auftretende Rostresistenz bei Kulturformen zeigte bisher, dass "ein einziges vollständig dominantes Gen (R) Rostresistenz bewirkte" (Sampson 1960, p. 216). Hier liegt der Gedanke einer Rückmutation oder Wiedergewinnung der Funktion eines Stillen Gens nahe. (Interessant ist in diesem Zusammenhang die Beobachtung von Voronova und Sefton 1986, dass die Expression einer neuentdeckten Tyrosinproteinkinase durch Insertion eines Retroviruspromotors um das 3-9fache gesteigert wird.) Über die genetischen Sequenzen und biochemischen Ursachen ist jedoch bei Antirrhinum noch nichts bekannt, so dass zu einer endgültigen Klärung der Frage noch eine erhebliche Forschungsarbeit notwendig ist.
Da viele Wildformen von Antirrhinum gegen Anfang der 1930er Jahre von den USA nach Europa eingeschleppten Antirrhinum-Rost resistent sind, ist offenbar bei den Kulturformen eine ursprünglich bei Antirrhinum generell gegebene Genfunktion verloren gegangen.
Weitere Beispiele für Resistenz durch dominante Gene bei Muona et al. 1982 und Nair et al. 1982.
Wiederholt sind auch Resistenzerscheinungen aufgrund von Rekombination beschrieben worden (z.B. Martens et al. 1981, Singh und Morton 1981 u.a.). Im Falle polygener Resistenz kann man sich die Situation mit Hilfe der obigen Rekombinationsquadrate (pp. 80, 122) und der Rekombination beim Anthozyansystem (p. 193) klarmachen. Einen interessanten Fall schildern uns Thukral et al. in ihrer Arbeit BIOCHEMICAL GENETIC BASIS OF DOWRY MILDEW RESISTANCE IN PEARL MILLET (1986, p. 648):
Phenolic content and the activities of peroxidase and polyphenol-oxydase were found to be related to degree of resistance. The immune genotype L 5 had the maximum, and the highly susceptible genotypes L 10 and A 7 had the minimum phenolic content, peroxidase activity and polyphenoloxidase activity.
Häufig sind jedoch die biochemischen Ursachen noch ungeklärt. Man könnte sich bei polygen bedingter Resistenz auch schwach nachteilige Veränderungen an mehreren Genloci vorstellen, deren Gesamteffekt die beobachtete Resistenz gegen z.B. Insektizide ist. Da wir bei den rezessiven Punktmutationen wiederholt gesehen haben, dass der Ausfall oder die verminderte bzw. vermehrte Produktion bestimmter Stoffe die Resistenz ermöglichte, kann dasselbe Prinzip natürlich auch auf polygener Ebene wirksam sein, da ja an der Bildung der meisten Verbindungen Genwirkketten beteiligt sind.
Durch Genamplifikation wird ebenfalls die vermehrte Produktion bestimmter Verbindungen ermöglicht. Fogel et al. haben dafür ein schönes Beispiel bei der Hefe bearbeitet. Sie schreiben 1983, p. 347 u.a.:
The CUP1 locus in yeast confers resistance to copper toxicity. We determined the molecular basis for copper resistance in three yeast strains, with differing degrees of resistance. Increased resistance to copper is associated with overproduction of a low molecular weight copper-binding Protein, copper-chelatin. Increased chelatin synthesis results from amplification of the CUP1r gene and increased synthesis of the copper inducible mRNA. The copper resistance level of a given strain correlates directly with the gene copy number.
Ich möchte auf die Amplifikations-Frage beim Thema Genduplikation zurückkommen und an dieser Stelle nur feststellen, dass weder neue komlexere Substanzen noch differenziertere Stoffwechselwege für die Resistenz notwendig sind, was praktisch auf alle bisher behandelten Fälle mit Ausnahme der Rückmutationen und Wieder-in-Betriebnahme von Stillen Genen zutrifft. Aber auch in den letzteren Fällen wird nichts prinzipiell Neues geschaffen, sondern es werden nur Schäden behoben: im ersteren Fall muss die notwendige spezifische DNA-Sequenz mit über 99prozentiger Genauigkeit noch vorliegen, damit eine Rückmutation im Bereich der Wahrscheinlichkeit liegt und im zweiten könnte der vollständigen Sequenz der deletierte Promotor durch Transposons wieder zugeordnet werden.
Der Ploidiegrad kann ebenfalls mit einer vermehrten Produktion bestimmter Verbindungen einhergehen, wie allgemein aus der Pflanzenzüchtung bekannt ist. Die Situation ist jedoch hier komplexer als bei der Beschränkung auf die Amplifikation bestimmter Gene, da hier die Wachstumsparameter des ganzen Organismus verändert sind. So sind z.B. diploide Rassen der Süßwasserschnecke Bulinus gegen den die Bilharziose verusachenden Trematoden Schistosoma haematobium resistent während die tetraploide Bulinus truncatus ein bedeutender Zwischenwirt dieses Parasiten ist.
Im allgemeinen ist die Fertilität experimentell produzierter Polyploider verringert (Details vgl. Gottschalk 1976, 1978).
Den Verlust der Salzresistenz mit zunehmendem Ploidiegrad bei Chenopodium in der Natur beschreiben Tanaka und Tanaka 1980, p. 266):
The halophytic Chenopodium glaucum was a diploid with 2n = 18. The semi-halophytic C. acuminatum var. japonicum was a tetraploid with 2n = 36. The non-halophytic C. album and C. album var. centrorubrum, on the other hand, were a hexaploid with 2n = 54. Presumably the euploidy has played a role of ecological differentiation in the four taxa.
Zur Zunahme von Kälteresistenz mit dem Polyploidiegrad vgl. die Diskussion bei Gottschalk 1976. Derselbe Verfasser stellt 1978/1984 p. 282 fest:
In der Frühzeit der Erforschung cytogeographischer Fragestellungen ist folgende Faustregel aufgestellt worden:
- je nördlicher das Florengebiet
- je extremer der Standort
- je höher die Lage des Areals
- umso höher der Anteil der Polyploiden
Sie mag in zahlreichen Einzelfällen stimmen, kann jedoch in dieser einfachen Form nicht verallgemeinert werden.
Unter Berücksichtigung neuerer Befunde wird heute die These vertreten:
- je jünger die Flora
- um so höher ihr Anteil an Polyploiden
Von einer allgemeinen Resistenzsteigerung durch Polyploidie kann demnach nicht die Rede sein.
Die Punkte 5 und 6 (p. 373) beziehen sich auf Bakterienresistenzen. Sie erklären vor allem die plötzlich auftretenden Vielfachresistenzen gegen Antibiotika, da hier mit Hilfe von Transposons und Plasmiden ein horizontaler Gentransfer zwischen verschiedenen Bakterienarten realisiert wird.
Mitsuhashi hat diesen Punkt 1982, p. V folgendermaßen zusammengefasst:
A mixed incubation of a small number of bacterial cells carrying transmissible drug resistance results in the rapid acquisition of this resistance by a majority of the other incubated cells. R factors are transferable among all species of the family Enterobacteriaceae, the Vibrio group, Pasteurella pestis, Bordetella bronchiseptica, and among the Pseudomonas group. These properties of the R factor, i.e., autonomous replication and its capability to transmit widely among various species of bacteria, are of great importance in public health and animal husbandry.
Durch den horizontalen Gentransfer werden bei Bakterien von einer Art zur anderen u.a. enzymatische Fähigkeiten vermittelt, wie beispielsweise die Übertragung des Kodes für das Penicillin-spaltende Enzym Penicillinase. Diese Fähigkeit dürfte allerdings so alt (oder fast so alt) wie die Interaktionen zwischen Pilzen und Bakterien und Bakterien untereinander sein. Der Transfer solcher DNA-Sequenzen wird sich im Laufe der Erdgeschichte millionenfach abgespielt haben. Bei nachlassender Selektion auf Resistenzfaktoren wird einige Zeit nach erfolgtem Gentransfer der Kode auch wieder millionenfach durch richtungslose Punktmutationen degeneriert sein. Die Bakterien, die unter anhaltenden Selektionsbedingungen den Kode aufrechterhielten, haben im Falle erneut einsetzender Selektion bei den anderen Bakterienarten den Kode dann wieder millionenfach vermittelt. Die Bakterien entwickeln sich dadurch nicht zu höheren Lebensformen. Sie bilden dadurch weder einen Zellkern noch entwickeln sie sich zu Vielzellern.
Wie wir schon im Detail diskutiert haben, ist die DNA zwar ein (absolut) notwendiger, aber kein hinreichender Grund für die Morphogenese (vgl. pp. 213-269 u. 325-327). Durch den horizontalen Gentransfer entstehen nicht einmal nach systematischen Kriterien neue Arten. Die Regel der Rekurrenten Variation gilt auch für die durch Plasmide vermittelten Sequenzen bei Bakterien.
Sollte ein vergleichbarer Gentransfer durch Retroviren bei Eukaryonten möglich sein, dann wären damit ebensowenig neue Formen zu erwarten.
Auf Details möchten wir beim Transposonthema zu sprechen kommen.
Den Punkt 7 (Selektion von Repair-Systemen) habe ich schon mit einem Beispiel der Ataxia telangiectasia erwähnt. Nicolas und Nigon berichten in ihrer Arbeit PRODUCTION OF PLASTIDIAL ANTIBIOTIC-RESISTANT MUTANTS BY ULTRAVIOLET IRRADIATION OF Euglena gracilis (1982, pp. 184 und 185):
Ultraviolett irradiation of Euglena gracilis drastically increases the proportion of green colonies resistant to chloroplast-inhibiting antibiotics. Some green colonies persist at high UV doses. These results might indicate the occurrence in the chloroplastic DNA of repair processes, of which one could be more error-prone.
...an error-prone repair, presumably acting on the ct DNA, might produce chloroplastic Ant r mutants.
Je schneller und fehlerhafter das Reparatursystem läuft, um so größer werden die Möglichkeiten der Bildung aberranter Substanzen und Stoffwechselwege (bzw. des Ausfalls funktionsfähiger Verbindungen) und um so geringer wird die Zahl der Ansatzpunkte gefährlicher Stoffklassen. Wie bedrohlich diese Methode für das Gesamtsystem des Organismus werden kann, ist mit der Ataxia telangiectasia schon veranschaulicht. Generell kann von Höherdifferenzierung bei den Resistenzerscheinungen nicht die Rede sein.
Kommen wir zum Punkt 8: Von Gressel stammt 1985 eine ausgezeichnete Studie zur Plastiden-gebundenen Herbizidresistenz von Kulturpflanzen, die unseren bisherigen Eindruck zu den Resistenzerscheinungen noch weiter vertieft. Im folgenden möchten wir die Hauptpunkte aus dieser Arbeit zitieren.
Nach Hinweis auf Haldanes Erkenntnis, dass sich die Kosten der Selektion meistens in einer herabgesetzten Fitness ausdrücken, schreibt er zu seinem Thema 1985, p. 490/491:
The case histories of herbicide resistance look pretty depressing vis a vis fitness (Tables 13, 14).
...The data in Tables 13 and 14A are not biased towards showing only cases where the susceptibles give greater yield. They are an exhaustive compilation. The near 20% greater yield found in the vast majority of S [Susceptible] biotypes is listed in the original articles as statistically "non-significant". Most scientists performing such experiments are not used to small differences and rarely use the required number of replicates. Haldane was right; there is a consistent loss of fitness upon selection for herbicide resistance.
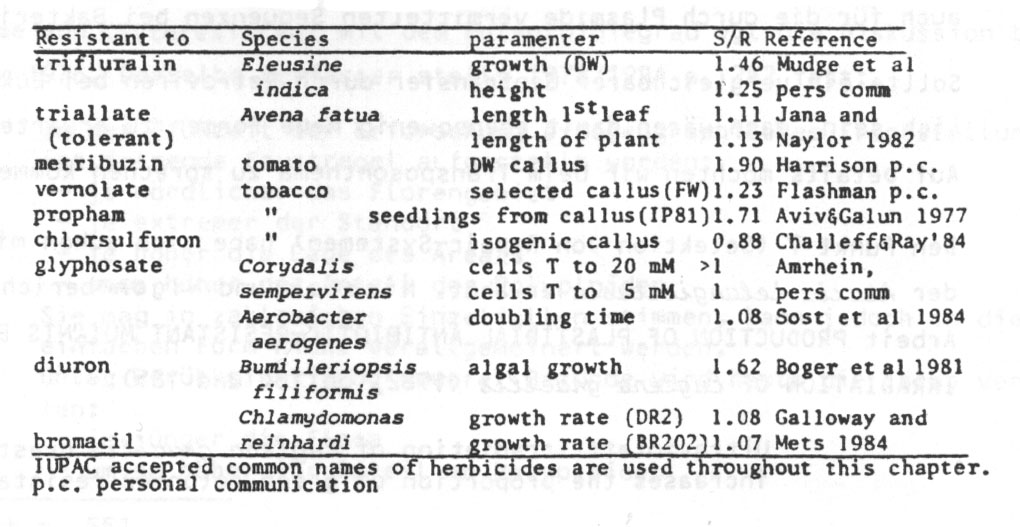
Sehen wir uns die Tabellen näher an (zu den genauer untersuchten Parametern wird das Verhältnis von nichtresistenten (S) zu resistenten (R) Linien ganz rechts angegeben).
Tabelle 13 (aus Gressel 1985): Productivity differences between herbicide R and S biotypes
Tabelle 14 (aus Gressel 1985): Decreased fitness of plastid inherited triazine resistance

Gressel kommentiert dazu nach einigen Bemerkungen zur Methodik der Fitnessprüfung pp. 491/492:
Competitive fitness has only been measured in triazine-resistant biotypes (Table 14B). If there were no differences in competitive fitness, an equal mixture of R and S biotype seeds should give rise to an equal mixture of seed at the end of the season. Almost all R biotypes had low fitnesses (Table 14B), lower than would have been predicted from the lower productivity of the same species (Table 14A). One of the few exceptions was Chenopodium strictum. A dose perusal of the data shows that this species grows at about half the rate of C. album under identical conditions; i.e. the factors lowering fitness were not the rate limiting factors. R and S biotypes should be tested under different environmental conditions to be truly judged. The fitness differential might be changed by water, light, temperature, seeding density, etc. Indeed, one of these parameters was partially checked with C. album. S plants grown separately at 25° C, produced more seed than the R. At 30° C there was no significant difference in seed production (Table 2A), but at 30° the growth rate of the (S) plants was half that of the rate at 25°.
Der Autor geht auch auf die biochemische Seite der Frage ein. Beim Mais scheint ein dominantes Gen für Glutathion-S-Transferase (GSH) zur Resistenz gegen s-Triazin eine besondere Rolle zu spielen, aber hier sind noch viele Probleme offen. Dieses Gen ist in den meisten Linien bereits vorhanden, so dass hier nicht einmal Rückmutationen für die Resistenz notwendig sind. Viele Studien haben sich mit der Triazin-Resistenz von Unkräutern beschäftigt, bei denen die Resistenz immer wieder auf der Plastidenebene aufgetreten ist. Gressel führt die Hauptpunkte p. 496 auf:
The most rapid difference is a 10 fold slower rate of decay of fluorescence measured microseconds after nano-second flashes of saturating laser light. This indicates a slower rate of transfer of electrons from reduced Q to B in photosystem II. Oxygen evolution was measured after short flashes of light spaced at different intervals in R and S Senecio plastids. Holt et al. (1983) conclude that their data "are consistent with a hypothesis" that the R plastids have more back-reactions from the reducing to the oxidizing side of PS II. This results in lower oxygen yield and increased "misses" and "double hits". The R plastids are only half as efficient in the Hill reactions as the S plastids at all light intensities (Table 14A). When C02 fixation, the process of ultimate importance in photosynthesis, is followed, the R leaves are much worse at lower intensities than they are at field levels of light (Holt et al. 1982).
Solche Funktionsverluste sind in voller Übereinstimmung mit der Fitnessabnahme. Das für die Atrazinbindung zuständige 32 kD Protein zeigt nach den bisherigen Untersuchungen nur die Substitution in einer Aminosäure (Serin zu Glyzin). Da das Protein stark konserviert ist, werden von einigen Molekulargenetikern für diese Frage keine weiteren Austausche erwartet, andere Überlegungen halten Substitutionen in weiteren Bereichen des Proteins jedoch auch für höhere Pflanzen für wahrscheinlich, nachdem bei Chlamydomonas Austausche bis zu vierzig Aminosäuren entfernt mit (wenn auch recht schwacher) Resistenz aufgetreten sind (Details bei Gressel). Verbesserte Fitness durch Resistenz erwartet beim Vergleich von R und S Linien ohne Herbizideinsatz niemand. Selbst bei starkem Herbizideinsatz sind resistente Unkräuter nur erschienen, wenn die Landwirte wiederholt Monokulturen ohne Fruchtwechsel anpflanzten und dabei immer wieder dasselbe Herbizid anwandten. "No cases of new weed resistance in fields have yet been reported where crop and herbicide rotations were used" (Gressel p. 490). Mit anderen Worten können sich die resistenten Genotypen mit ihrem herabgesetzten Informations- und Funktionsniveau erst nach solchen repetitiven Vernichtungsfeldzügen durchsetzen. Für die Populationen ist das unter solchen Voraussetzungen die einzige Möglichkeit zu überleben. Wird jedoch nach einiger Zeit der Herbizideinsatz aufgehoben, so kehren die Populationen (wohl durch Zuwanderung und/oder Rückmutation und Transposonaktivität) zu ihrem ursprünglichen Funktionsniveau zurück und die Resistenz ist wieder aufgehoben. Dafür gibt es aus der gesamten Pestizidforschung unzählige Beispiele und die Regel gilt sowohl für die Pflanzen- als auch die Tierwelt (z.B. Verlust der DDT-Resistenz vieler Tierpopulationen), - was im Falle von Rückmutationen und Transposons ein weiteres Beispiel für die Regel der Rekurrenten Variation ist.
Zum letzteren Punkt sei noch hervorgehoben, dass auch bei der beschriebenen Herbizidresistenz die Genotypen in der Regel nicht erst neu auftreten, sondern in den Populationen schon vorhanden sind. Nach den Untersuchungen von Gasquez und Compoint 1981 repräsentiert eine resistente Linie jeweils eine der vielen Isozymvarianten der Population an dem jeweiligen Areal. Weitere Beispiele für die Regel der Rekurrenten Variation bei Grupp und Annear 1981, Singh und Pandey 1982, Csépló und Maloga 1984.
Zusammenfassend können wir feststellen, dass die Resistenzphänomene in guter Übereinstimmung mit der Regel der Rekurrenten Variation und dem mutativen Abbau von Information, Struktur und Funktion sind.
________________________
NACHTRÄGE ZU DEN SEITEN 378 (OBEN) UND 383 (RESISTENZERSCHEINUNGEN)
Mouchès, C., N. Pasteur, J.B. Bergé, O. Hyrien, M. Raymond, B.R. de Saint Vincent, M. de Silvestri und G.P. Georghiou (1986): AMPLIFICATION OF AN ESTERASE GENE IS RESPONSIBLE FOR INSECTICIDE RESISTANCE IN A CALIFORNIA CULEX MOSQUITO. Science 233, 778 - 780.
P. 778: Adults of the Tem-R strain were found to posses at least 250 times more copies of the gene than adults of a susceptible strain (S-Lab). The finding that selection by pesticides may result in the amplification of genes encoding enzymes in whole, normally developed reproducing insects emphasizes the biological importance of this mechanism and opens new areas of investigation in pesticide resistance management.
Ähnliches ist auch bei Pflanzen gefunden worden:
Klee, H.J., Y.M. Muskopf und C.S. Gasser (1987): CLONING OF AN ARABIDOPSIS THALIANA GENE ENCODING 5-ENOLPYRUVYLSHIKIMATE-3-PHOSPHATE SYNTHASE: SEQUENCE ANALYSIS AND MANIPULATION TO OBTAIN GLYPHOSATE-TOLERANT PLANTS. Mol. Gen. Genet. 210, 437 - 442
P. 437: We have fused the gene to cauliflower mosaic virus 35 S promoter and reintroduced the chimeric gene into Arabidopsis. The resultant overproduction of EPSPs leads to glyphosate tolerance in transformed callus and plants.
Resistenz durch rezessive Gene:
Jorgensen, J.H. (1983): EXPERIENCE AND CONCLUSIONS FROM THE WORK AT RISO ON INDUCED MUTATIONS FOR POWDERY MILDEW RESISTANCE IN BARLEY. IN: INDUCED MUTATIONS FOR DISEASE RESISTANCE IN CROP PLANTS II. IAEA (1983, pp. 73 - 87).
P. 75: The...10 independently induced genes were tested for inheritance and for allelism by appropriate crosses and by testing with pathogen culture A-6 (290). In all 10 cases the resistance was conferred by a single recessive gene. The 10 genes were non-complementing alleles in one locus. (Dazu ist noch eine Spontanmutation im selben Locus beschrieben worden.)
Ein (weiteres) schönes Beispiel für Resistenz durch Genfunktionsausfall:
Handtke, K. (1987): SELECTION PROCEDURE FOR DEREGULATED IRON TRANSPORT MUTANTS (fur) IN ESCHERICHIA COLI K12: fur NOT ONLY AFFECTS IRON METABOLISM. Mol. Gen. Genet. 210, 135 - 139.
P. 135: ...it could be shown that nearly all the mutants constitutive in the expression of the operon fusion fiu: [lamda]p lacMu were only defective in fur. ...Most of the fur mutants isolated (80%) were not able to grow on succinate, fumarate or acetate. After transformation with a fur+ plasmid all 39 mutants tested were able to grow on succinate.
P. 138: Screening for manganese-resistant mutants selected, with high incidence, mutants in the central regulatory gene of iron metabolism, fur. ...Whatever the mechanism of manganese action is, the constitutive mutants were not repressed by Mn2+ and this allowed the cells to grow.
___________________
Nachtrag (2001): Vgl. weiter die ausführliche
Diskussion samt
systematischer Widerlegung unzutreffender
Einwände und weit verbreiteter Irrtümer zum Thema der evolutionistischen Bedeutung der Antibiotikaresistenzen (2000/2001).