5. DER PALÄONTOLOGISCHE ARTBEGRIFF
Auf der Seite 47 haben wir festgestellt, dass der paläontologische Artbegriff praktisch mit dem morphologischen gleichzusetzen ist. Der schon erwähnte Paläontologe Clarkson schreibt zu diesem Punkt (1979, p. 7) [vgl. auch 1986, p. 6]:
The fundamental unit of taxonomy is the species. Animal species...are groups of individuals that generally look like each other and can interbreed together to produce offspring of the same kind. They cannot interbreed with other species. Since it is reproductive isolation alone that defines species it is only really possible to distinguish closely related species if their breeding habits are known. Of all the described 'species' of living animals, however, only about a sixth are 'good', or properly defined, species. Information upon the productive preference of the other five-sixth of all naturally occurring animal populations is just not documented. The differentiation of most living and all fossil species therefore has to be based upon other and technically less valid criteria.
Of these by far the most important, especially in paleontology, is morphology, the science of form, for most natural species tend to be composed of individuals of similar enough external appearance to be identifiable as of the same kind.
Da rund 5/6 aller heute erfassten Tierarten sowieso nur morphologisch definiert sind, unterscheidet sich der paläontologische Artbegriff nach Clarksons Auffassung offensichtlich nicht wesentlich vom neontologischen, zumal die meisten natürlichen Arten aus ähnlichen Individuen bestehen.
Kull beurteilt die Situation hingegen (1979, p. 243) wie folgt:
Fossilien sind immer nur Reste, die kleinere oder größere Teile der Morphologie und Anatomie der Lebewesen erkennen lassen. Die Art ist aber populationsbiologisch definiert (vgl. 3.5.1.). Wenn man bei heutigen Formen nicht sicher weiß, ob sie zu einer Art oder zu zwei getrennten Arten gehören, kann man dies durch Kreuzungsversuche klären. Bei fossilen Organismen ist dies nicht möglich. Da außerdem an Fossilien oft auch für die Artanerkennung wichtige Merkmale nicht erhalten sind, ist der Artbegriff bei Fossilien (paläontologischer Artbegriff) vom biologischen Artbegriff grundsätzlich verschieden. (Kursiv vom Verfasser.)
[Vgl. auch G. Laurent 1987, p. 141 ("Cuvier donne une définition biologique de 1'espèce, fondée sur la génération. Elle est évidemment inutitisable en Paléontologie.") unter Hinweis auf einen Beitrag von Mahé und Devillers 1982. Siehe weiter auch Thomas und Spicer 1987, p. 72 ("Strictly speaking a palaeobotanical species is not equatable to a species of a living organism." Unter günstigen Voraussetzungen wird die Kluft jedoch kleiner).]
Nun haben Paläontologen wie Dacqué, Beurlen, Schindewolf, Kuhn, Gould, Eldredge, Stanley und viele andere darauf hingewiesen, dass das paläontologische Material zwar an manchen Stellen sehr lückenhaft ist ('Würmer', Quallen und andere skelettlose Organismen), dass auf der anderen Seite jedoch in vielen Tiergruppen ein "geradezu erdrückendes fossiles Material vorliegt (Foraminiferen, Korallen, Brachiopoden, Moostiere, Cephalopoden, Ostracoden, Trilobiten usw.).." (Kuhn 1965, p. 5, ähnlich 1981). Mit solchem bestens erhaltenen Material, bei dem häufig für jede Form und Formabweichung buchstäblich Millionen von Individuen vorliegen, kann man sehr wohl detaillierte morphologisch-anatomische Studien treiben und gründliche Überlegungen zum Artbegriff vornehmen.
Die für die Paläontologie zu konstatierende grundsätzliche Unmöglichkeit, Fragen der Artzugehörigkeit durch Kreuzungsversuche zu klären, hat jedoch bei vielen Paläontologen zu einem Verzicht auf den Anschluss an neuere Bemühungen um eine naturwissenschaftlich-genetische Klärung des Artbegriffs geführt, und nahezu sämtliche von Paläontologen beschriebene Arten sind Arten im oben zitierten morphologischen Sinne: jede nur denkbare Formabweichung erhielt (und erhält) in der Regel eigenen Artstatus, bei 'größeren' Unterschieden (was immer man für groß genug gehalten hat und hält) wird auch mit dem Gattungsbegriff nicht gespart. Man braucht sich nur einmal vorzustellen, wieviele neue Arten und Gattungen man nach paläontologischer Praxis allein bei den rund 450 Rassen unseres Haushundes (Canis lupus familiaris) aufstellen könnte, wenn man die Skelett- und Größenunterschiede zwischen den Rassen zum Maßstab für den Art- und Gattungsbegriff machen würde, - oder wie viele Hunderte und Tausende neuer Arten man nach solcher systematischen Praxis ganz allgemein bei unseren Haustieren und Kulturpflanzen beschreiben könnte.
Inwieweit kann uns hier der Artbegriff der Synthetischen Evolutionstheorie weiterhelfen? Seine generelle Unzulänglichkeit wird sowohl für die Pflanzen-und Tiergeographie als auch die Paläontologie u.a. von Ax hervorgehoben. Nach Zitat der Mayrschen Artdefinition (vgl. p. 38 unten) kommentiert er (1984, pp. 23/24):
In dieser Kennzeichnung wird die Aufrechterhaltung des gemeinsamen Genpools durch Kreuzungen und Reproduktionsbarrieren als zentrale Komponente der Kohäsion zwischen den Individuen einer Art herausgestellt. Trotzdem bleibt das biologische Artkonzept für die Interpretation der Art als Grundeinheit der Evolution unzulänglich.
Ein wesentlicher Einwand betrifft den Mangel einer objektivierbaren raum-zeitlichen Begrenzung der Art. In Konsequenz der Definition kann sich das biologische Artkonzept nur auf zeitlich und räumlich koexistierende Populationen erstrecken, auf synchrone und sympatrische Populationen verschiedener Arten also, in denen Kreuzungen aktuell möglich sind und zwischen denen reproduktive Isolationsmechanismen de facto wirksam werden. Engagierte Verfechter der Definition unterstreichen selbst den nicht-dimensionalen Charakter des biologischen Artkonzeptes: "As one progresses geographically and chronologically further and further away from a single point, the species distinctiveness beeomes more and more vague" (BOCK 1979, p. 28).
In der räumlichen Dimension bleibt es im biologischen Artkonzept unentscheidbar, ob phänetisch identische, allopatrische Populationen, die in der Folge ökogeographischer Barrieren reproduktiv voneinander getrennt sind und zwischen denen die Potenz zur Kreuzung und Erzeugung fertiler Nachkommen nicht nachweisbar ist, Angehörige einer Art sind oder nicht.
Schwerer wiegt jedoch die Frage nach der Begrenzung der Art in der Zeit. Über die erstaunliche Behauptung, eine aus einer Serie sich miteinander kreuzender Populationen bestehende Art sei heute einfach nicht dieselbe wie ihr Vorfahre vor 100 Generationen, kommt BOCK (1979, p. 29) zu dem Schluß: "Because the biological species concept is a nondimensional one, it is not possible to speak of the age of a species or the origin of a species, or of the life and death of a species."
Eindeutiger als in diesem Satz kann die Untauglichkeit eines nicht-dimensionalen Artkonzeptes zur Charakterisierung von Arten als reale Einheiten der Natur wohl kaum zum Ausdruck kommen. Sofern eine Art im Verlauf von 100 oder beliebig mehr Generationen sich nicht in mindestens zwei reproduktiv getrennte Einheiten aufspaltet, gehören die Populationen, welche vor einer entsprechenden Zeit existiert haben, und die heute lebenden Populationen selbstverständlich zu ein und derselben Art, - und zwar schlicht deshalb, weil sie als Glieder einer ununterbrochenen Ahstammungslinie miteinander in einem kontinuierlichen genealogischen Zusammenhang stehen. Wohl niemand kann ernsthaft bestreiten, daß die Populationen des heutigen Menschen wie die Populationen, die vor 100 Generationen oder einigen tausend Jahren die Erde bevölkerten, gleichermaßen zur Art Homo sapiens gehören. Für das Problem der zeitlichen Limitierung ist es dabei absolut unerheblich, ob in den durch Fortpflanzung in der Zeit zusammengehaltenen Linien von Arten evolutive Neuheiten entstehen oder nicht, ob also mit anderen Worten das Erscheinungsbild sukzedierender Populationen von Arten Veränderungen erfährt oder konstant bleibt.
Dass der Hauptgrund für die allzu eng gezogene Artabgrenzung des 'biologischen' Artbegriffs in der Zielsetzung liegt, (geographisch) Spezies in statu nascendi zu demonstrieren (und mit solcher Artentstehung den Modus des Ursprungs aller Lebensformen zu implizieren), haben wir oben schon detailliert aufgezeigt und wiederholt betont. Es ist geradezu der Sinn dieses Artbegriffs, dass die Spezies an Deutlichkeit und Klarheit verlieren, je weiter man sich geographisch und chronologisch von einem Punkt entfernt (vgl. das Zitat nach Bock, vorherige Seite). Die Spezies "sollen" ja kontinuierlich in andere Spezies übergehen. Unter Voraussetzung dieser Kontinuitätsidee gibt es natürlich auch keine Speziesgrenzen in der Zeit, und eine heutige Art braucht nicht mehr identisch mit der Art ihrer Vorfahren vor 100 Generationen zu sein. Die einfache Projektion unserer Rekombinationsquadrate (pp. 80, 122, 125, 181, 183) in die Zeit veranschaulicht uns diesen Punkt: Durch Abbau und Rekombination genetischer Strukturen können zwischen zwei durch hundert Generationen voneinander getrennte Populationen vielfache prä- und postzygotische Isolationsbarrieren entstanden sein (deren Nachweis allerdings nur möglich wäre, wenn man diese beiden Populationen wieder auf die gleiche Zeitebene bringen könnte).
Willmann lehnt jedoch (1985, pp. 125-128, 169-171) einen solchen Ansatz grundsätzlich ab, denn "das Kriterium der Fortpflanzungsisolation innerhalb einer evolutiven Linie entlang der Zeitachse anzuwenden, bedeutet, unvereinbare Bezugssysteme miteinander zu vermischen. ...Allochrone Populationen gehören...nie zu ein und derselben Fortpflanzungsgemeinschaft'' (pp. 125/126). Wir kommen darauf noch zurück.
Ax setzt an die Stelle des neodarwinistischen Artbegriffs "das evolutionäre Artkonzept", welches solche Probleme "durch Berücksichtigung der Dimensionen Raum und Zeit, Abdeckung des Spektrums unterschiedlicher Reproduktionsmodi sowie Kompatibilität mit allen Modellen der Artbildung" überwindet und definiert mit Simpson (1961) in der revidierten Fassung von Wiley (1978, 1981) die Art folgendermaßen (1984, p. 24):
An evolutionary species is a single lineage of ancestor-descendant populations which maintains its identity from other such lineages and which has its own evolutionary tendencies and historical fate.
- wobei 'lineage' eine Serie von Populationen mit einer gemeinsamen Abstammungsgeschichte bezeichnet.
Man sieht sofort, dass dieses Artkonzept wesentlich umfassender als das der Synthetischen Evolutionstheorie ist.
Wir wollen auf Einzelheiten dieses Artkonzepts zu sprechen kommen, indem wir uns den Beispielen Willmanns zuwenden. Dieser Paläontologe hat unter Einbeziehung der kladistischen Methodik versucht, den 'biologischen' Artbegriff in die Paläontologie einzuführen. Wie ich jedoch schon betont habe, geht Willmanns Biospezieskonzept über das neodarwinistische insofern hinaus, als die Grenzziehung zwischen den Arten durch die "absolute Fortpflanzungsisolation" definiert ist (vgl. Willmann 1985, pp. 48-50) (kritische Anmerkungen s.u.). Wiederholt hebt er hervor, dass das 'Biospezies-Konzept definitionsgemäß kein merkmalsbezogenes Artkonzept ist' (pp. 132-133), was zum einen die Unabhängigkeit vom morphologischen Artbegriff bedeutet, zum anderen jedoch die Anerkennung von genetisch absolut voneinander getrennten Zwillingsarten heißt, - ein Punkt, der uns nachher noch beschäftigen soll.
Trotz mancher Probleme (vgl. p. 300f.) ist auch dieser Ansatz insofern ein großer Schritt in die richtige Richtung, als mit diesem Artbegriff Kriterien vorgelegt werden, die bei konsequenter Anwendung der uferlosen Speziesmacherei in der Paläontologie ein Ende bereiten würden. Für eine Revision in der Paläontologie durch sein Spezieskonzept liefert der Autor eine Reihe überzeugender Beispiele, von denen hier ein Überblick gegeben werden soll.
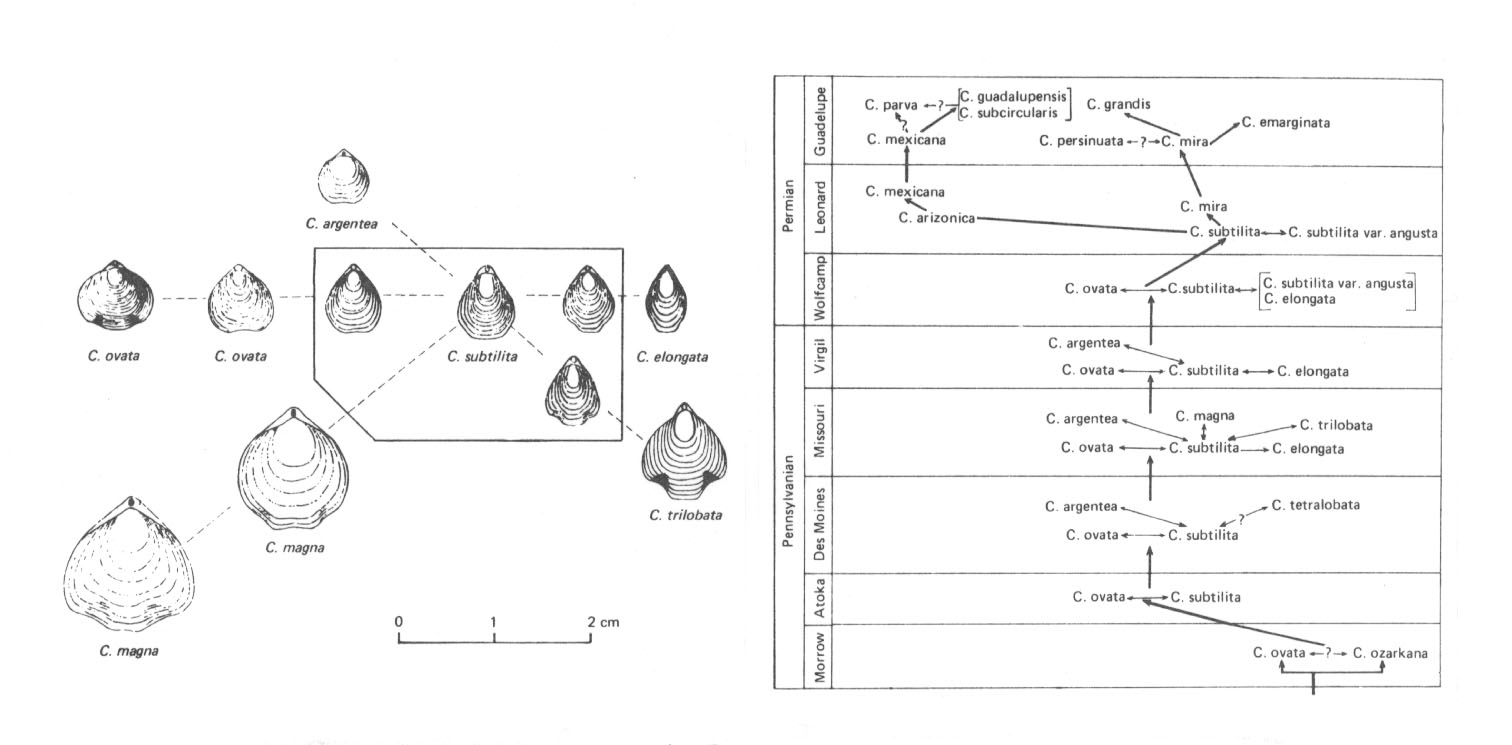
Bei Composita subtilita (Brachiopoda) wurden 1964 noch 17 verschiedene Arten durch die angenommenen Zeithorizonte unterschieden. "Die typologische Methodik erforderte in diesem Schema die Berücksichtigung von bis zu sechs gleichzeitigen Spezies. Tatsächlich handelt es sich im wesentlichen (vom Unterkarbon bis in die Leonard-Stufe) um den evolutiven Wandel einer einzigen Art" (pp. 88 - 90). Wir sehen an diesem Beispiel bereits die Anwendung des oben nach Wiley zitierten Artbegriffs als "a single lineage of ancestor-descendent populations which maintains its identity from other such lineages and which has its own evolutionary tendencies and historical fate."
Abb. 45: Links: die morphologischen Arten des Brachiopoden Composita subtilita. - Diese Arten werden in der Abbildung rechts in den verschiedenen Erdschichten oft nebeneinander aufgeführt. Die mit nur einer Spitze versehenen Pfeile sollen die Abstammungsverhältnisse angeben, die Pfeile mit zwei Spitzen weisen auf Beziehungen der Rekombinanten innerhalb der Population hin. Bis zu sechs gleichzeitigen Spezies werden nach dem morphologischen Artbegriff in das Schema eingetragen. "Tatsächlich handelt es sich im wesentlichen (vom unteren Oberkarbon bis in die Leonard-Stufe) urn den evolutiven Wandel einer einzigen Art." Wie sieht es jedoch mit den Formen der permischen Guadelupe-Stufe aus? Gemessen an der innerartlichen Variabilität heutiger Mollusken dürften diese Formen ebenfalls zu Composita subtilita gehören.
Aus Willmann 1985 nach Grinnel und Andrews. Verkleinert.
Was hier von Willmann als "evolutiver Wandel" bezeichnet wird, dürfte mit Mendelscher Rekombinatorik und Genökologie bereits voll zu erklären sein (Composita subtilita wird immerhin von der oberkarbonischen Atoka-Stufe bis zur permischen Leonard-Stufe verzeichnet - in geologischer Zeitrechnung eine Konstanz von über 40 Millionen Jahren).
Zur ganz erstaunlichen, allein durch Umweltfaktoren bedingten Variabilität (Modifikationen) beim nächstverwandten Stamm der Mollusken vgl. z.B. Arthus 1982, Burla und Stahel 1983 und Burla 1984.
Arthus bemerkt in seiner Arbeit CONTROL OF SHELL SHAPE IN Lymnaea stagnalis 1982, p. 153:
...the large difference in shape observed between the populations in the wild was completely due to direct environmental effects on the phenotype, or any genetic component of the variation was so small as to be undetectable by the method employed. It is noted that this result cautions against acceptance of the evolutionary inferences that are often drawn from studies of phenotypic variation in shell shape where these are unaccompanied by demonstration of an inherited component of the variation.
Zurück zu Willmann:
Als weitere Beispiele werden der Seeigel Micraster, die Ammonoideen Leymeriella und Ermoceras, die Koralle Zaphrentis delanonei aus dem englischen Unterkarbon, die Foraminifere Conorotatites, die jurassischen Ammoniten der einen ausgeprägten Geschlechtsdimorphismus aufweisenden Gattung Kosmoceras und die Foraminifere Neogloboquadrina diskutiert (pp. 93-95, 98/99, 100-104, 108, 137-140). In all diesen (und vielen weiteren) Fällen wurden Formen als eigene Arten beschrieben, die nach den vorliegenden paläontologischen Daten eine Fortpflanzungs- und Abstammungsgemeinschaft in Raum und Zeit bildeten und die, gemessen an den Merkmalsdivergenzen innerhalb heutiger Arten, sehr wahrscheinlich eine einzige Mendelpopulation bildeten.
Trotz stark unterschiedlicher Bewertung des neodarwinistischen Ansatzes (Willmann empfiehlt in seinem Vorwort 1985 Mayrs Werk ARTBEGRIFF UND EVOLUTION (1967) als grundlegende Ergänzung zu seinen Ausführungen), ist beiden Autoren gemeinsam, dass sie die Aufspaltung von einer Art in zwei reproduktiv voneinander isolierte Arten zum Angelpunkt ihrer Artdefinition machen (kladistischer Ansatz; Hennig 1950, 1966). Willmann schreibt (p. 131): "Die Entstehung von Biospezies besteht darin, daß zwei aus einer gemeinsamen Stammart hervorgehende Populationen im Verhältnis zueinander reproduktiv isoliert werden. ...Eine Art ist vor allem in Relation zu ihrem nächstverwandten Taxon eine Art. Daher sind die Populationen als neue Arten und die Stammart als erloschen aufzufassen." Speziation heißt immer Artaufspaltung einer Stammart in Tochterarten. "Eine Art ist der evolutive Abschnitt zwischen zwei Speziationen oder aber zwischen einer Speziation und dem Zeitpunkt ihres nachkommenlosen Aussterbens" (p. 133). Ebenso nennt Ax (1984, p. 29) die 'Entstehung neuer Arten durch Spaltung einer existierenden Art' den "entscheidenden Prozeß der Speziation". Zur Verdeutlichung seien noch einige weitere Sätze von Willmann zitiert (1985, pp. 118, 119, 121):
Eine Art entsteht als selbständige Fortpflanzungsgemeinschaft bei der Aufspaltung ihrer Mutterart, und die endet, sobald sie sich selbst in Tochterarten aufspaltet. Dieser Anfang und dieses Ende bilden die natürlichen Grenzen der Art im Zeitverlauf.
...Speziationsereignisse bilden im Zeitablauf die objektiven Artgrenzen. Damit ist der Gültigkeitsbereich des Biospezies-Konzeptes in die Zeit erweitert.
...Eine Art hört also in jenem Augenblick auf zu existieren, in dem die Nachkommen einer Fortpflanzungsgemeinschaft zwei Gruppen von Populationen bilden, die reproduktiv voneinander isoliert sind.
...Nach dem konsequenten Biospezies-Konzept entstehen zwei neue Arten ganz eindeutig zu jenem Zeitpunkt, zu dem reproduktive Isolationsmechanismen voll wirksam werden. Vor diesem Augenblick besteht nur eine Art, so weit vorangeschritten die Aufspaltung auch sein mag.
Ohne Aufspaltungsprozess hingegen bedeuten für diesen Artbegriff durch Transposons, Mutation und Rekombination bedingte anatomische und physiologische Veränderung (Schwerpunktverlagerungen) in einer Linie gar nichts, selbst wenn zahlreiche Indizien darauf hinweisen, dass solche Unterschiede bei gegenwärtigen Populationen eine absolute Isolationsbarriere bedingen würden. "In ein und dieselbe Spezies müssen zwei allochrone Populationen derselben evolutiven Linie auch dann gestellt werden, wenn z.B. aus Gründen der Konstruktion ausgeschlossen werden kann, daß ihre Vertreter sich erfolgreich verpaaren konnten ("wenn sie gleichzeitig gelebt hätten"). So etwas wäre z.B. denkbar bei einer beträchtlichen Größenzu- oder -abnahme innerhalb einer Formenreihe oder bei einem tiefgreifenden Umbau der Genitalarmaturen. In solchen Fällen können Ausgangs- und Endform einer evolutiven Linie so stark voneinander abweichen, daß, hätte es sich um gleichzeitige Populationen gehandelt, eine mechanische Fortpflanzungsisolation wirksam gewesen wäre. Daß solche Populationen eine einzige Biospezies bilden, steht aber nicht im Widerspruch zum Kriterium der Fortpflanzungsbeziehungen im biologischen Artkonzept, weil sie nicht gleichalt sind. Vergleichbar ist diese Situation mit den schon besprochenen "ring-species". Hier können sogar Populationen eine Biospezies bilden, die nachweislich reproduktiv voneinander isoliert sind." Durch "Artumwandlung" oder "Artabwandlung" nicht aufspaltender Linien entstehen keine neuen Biospezies. "Alle fossilen Taxa, die nur über diesen Vorgang entstanden sind, sind im Vergleich zu anderen Transformationsstufen derselben evolutiven Linie keine Arten im Sinne des biologischen Spezieskonzeptes. Evolutive Arten, wie lang und welchem Wandel auch immer unterworfen, gehören einer einzigen Art an, solange keine phylogenetische Aufspaltung erfolgt. Eine Art wird also zu verschiedenen Zeiten ein sehr verschiedenes Aussehen zeigen können" (Willmann 1985, pp. 128/129).
Ax vertritt die gleiche Auffassung, wenn er (1984, p. 27) schreibt:
Die weit verbreitete Auffassung über die Transformation von Arten in der Zeit (Artumwandlung, phyletische Evolution, phyletische Speziation) geht von dem Gedanken aus, eine Art könne sich über längere Zeiträume hinweg ohne irgendeinen Spaltungsprozeß in eine zweite, dritte, vierte...Art verwandeln.
...Es bedarf wohl nur weniger Worte, um die Unvereinbarkeit der Konzeption derartiger "Sukzessions-Arten" (successional species) mit dem evolutionären Artkonzept offenzulegen. In einer direkten Linie von Populationen, welche durch den Prozeß der Fortpflanzung kontinuierlich verbunden bleiben, mögen beliebig viele evolutive Veränderungen die genetische Information und damit das Merkmalsmuster in aufeinanderfolgenden Generationen beliebig stark verwandeln; ohne einen Spaltungsprozeß bleiben die einzelnen Individuen dieser Linie logischerweise die Glieder einer einzigen evolutionären Einheit. Die Transformation des Genoms in der Zeit führt nicht zur Umwandlung einer Art in eine andere, neue Art, sondern lediglich zum Wechsel in der Erscheinung zeitlich gestaffelter Generationen ein und derselben Spezies. ...Das Modell der Transformation von Arten repräsentiert keinen Speziationsmodus der Natur; es ist das artifizielle Produkt einer fehlerhaften Gedankenführung.
In diesen Zitaten wurde von "evolutivem Wandel", "evolutiver Linie", "beliebig vielen evolutiven Veränderungen", von der "Transformation des Genoms in der Zeit" etc. gesprochen, so dass man fast den Eindruck erhält, als ob sich tiefgreifende Veränderungen und Neuerungen bei den verschiedensten Linien paläontologisch am laufenden Band dokumentieren ließen. Wir möchten an dieser Stelle darauf hinweisen, dass mit solchen Begriffen, einschließlich des Zentralbegriffs dieses Artkonzepts, der Artaufspaltung, wie beim Neodarwinismus wieder der Modus des Ursprungs aller Lebensformen impliziert wird. In der Formulierung von Ax, dass "beliebig viele evolutive Veränderungen die genetische Information und damit das Merkmalsmuster in aufeinanderfolgenden Generationen beliebig stark verwandeln mögen" etc. werden die evolutiven Veränderungen der 'Verwandlung' der genetischen Information sogar übergeordnet ("die Evolution" erscheint in diesen und vielen anderen Arbeiten häufig als übergeordnetes Prinzip, das 'Ding an sich', das Leitmotiv, welchem sich alle anderen Gedankengänge unterzuordnen haben), so dass hier Ursache und Wirkung miteinander verwechselt werden. Es sind doch nicht die evolutiven Veränderungen, die die genetische Information verwandeln, sondern es sind die genetischen Möglichkeiten und Grenzen der Veränderung, die über 'die Evolution' entscheiden.
Um die Situation in der Paläontologie kurz und bündig zu kennzeichnen, sei ein nach wie vor treffendes Wort Schindewolfs zitiert (ähnlich Gould und Eldredge 1977, Gould 1983, Stanley 1979, 1984). Er schreibt 1965, pp. 85/86:
Nach darwinistischer Vorstellung sollen geringfügige Rassenunterschiede sich allmählich zu Artmerkmalen verstärken und diese dann durch Addition immer neuer kleiner Abänderungen zu Gattungs-, Familienunterschieden und so weiter werden. Die Formen-Mannigfaltigkeit müßte alsdann gegen Ende der einzelnen Stämme zunehmen; dort wäre die größte Fülle von Ordnungen, Familien und Gattungen, das heißt von Unterschieden höheren Grades zu erwarten. Das Gegenteil ist der Fall.
Ein neuer Bauplan von dem systematischen Range etwa einer Klasse oder Ordnung erscheint gewöhnlich völlig unvermittelt auf der Bildfläche, ohne lange Reihen von Bindegliedern, die uns eine allmähliche Herausgestaltung aus einer anderen, seine Wurzel bildenden Ordnung vor Augen führen würden. (Hervorhebungen im Schriftbild von mir.)
Aufgrund dieser in der Regel auch für die Familien und Gattungen zutreffenden Tatsache hat sich die punktualistische Schule in der Paläontologie (vgl. die oben erwähnten Paläontologen) von der Idee der kontinuierlichen Entwicklung abgewandt und postuliert statt dessen in Annäherung an Goldschmidt (vgl. z.B. Gould 1980) eine für die angenommenen entscheidenden Übergangsphasen stark beschleunigte Entwicklung in kleinen Populationen, die in den meisten Fällen paläontologisch nicht zu erfassen seien. An anderer Stelle möchten wir auf diese Probleme noch einmal zurückkommen (vgl. pp. 314-323) und jetzt nur einmal festhalten, dass die in den obigen Zitaten von Willmann und Ax implizierte (Gesamt-)Evolution aus den paläontologischen Daten nicht induktiv abgeleitet werden kann.