
DIE FRAGE NACH DER ANWENDUNG DES BIOSPEZIESKONZEPTS IN DER PALÄNTOLOGISCHEN PRAXIS
Wie sieht es nun mit der Anwendung des durch Artaufspaltung und absolute reproduktive Isolationsbarriere definierten Artkonzepts in der paläontologischen Praxis aus? Willmann schreibt, dass "wir den Zeitpunkt, wann die reproduktive Isolation erfolgt ist, an fossilen Organismen niemals ermitteln können" (p. 152) und führt das für die geographische und die sympatrische Speziation aus. Er fügt jedoch hinzu (ebenfalls p. 152):
Diese Konzession an die Praxis berührt die vorstehend erläuterte Theorie über die Position der Artgrenzen nicht. Wenn wir uns ein Speziationsereignis als einfache Verzweigung vorstellen..., dann ist es ohne tiefgreifende Bedeutung, ob wir den "taxonomischen Verzweigungspunkt" dort legen, wo sich zwei Populationen geographisch endgültig trennen oder etwas später - im Zeitpunkt der biologischen Isolation.
Obwohl auch nach Ax rezente und ausgestorbene Organismen systematisch grundsätzlich gleich behandelt werden sollten, hebt auch er die mit dem paläontologischen Material verbundenen Schwierigkeiten hervor und meint u.a. (p. 210):
Die Probleme beginnen schon mit der Erfassung und Abgrenzung evolutionärer Arten in der fossilen Überlieferung. Es gibt grundsätzlich keinen objektivierbaren Maßstab, um verschiedene individuelle Fossilien als Angehörige einer bestimmten singulären Art ansprechen oder aber als Repräsentanten reproduktiv getrennter Arten ausweisen zu können. Selbstverständlich wird man eine hohe Übereinstinimung oder Identinät in allen fossilisationsfähigen Strukturen als Indizien für die Zugehörigkeit verschiedener Individuen zu einer Art bewerten. Ebenso selbstverständlich wird man signifikante Unterschiede im Merkmalsmuster verschiedener Fossilien als Ausdruck einer einstmaligen reproduktiven Isolation deuten und die entsprechenden Individuen als Vertreter verschiedener evolutionärer Arten hypothetisieren. Da uns aber mit seltenen Ausnahmen - z.B. Einbettung viviparer Ichthyosauria mit Jungtieren im Mutterleib - der Fortpflanzungszusammenhang zwischen fossil überlieferten Individuen verschlossen bleibt, behalten die in diesem Bereich möglichen und notwendigen Entscheidungen selbst dann den Charakter subjektiver Einschätzungen, wenn überaus ähnliche fossile Fragmente an einem bestimmten Ort aus ein und demselben Zeithorizont geborgen wurden. Die hohe Irrtumsquote, die wir bei Einschätzung der Zahl ausgestorbener Arten in Rechnung stellen müssen, tangiert indes in keiner Weise die Validität des evolutionären Artkonzeptes und die Legitimität seiner Ausdehnung auf die fossile Überlieferung. Aus der erzwungenen Beschränkung auf Analysen und Vergleich fossilisationsfähiger Strukturmerkmale und der an sie gebundenen Verstärkung der Unzulänglichkeiten in unseren Systemen von Beobachtung und Methode (S. 31) kann nicht die Spur einer Berechtigung abgeleitet werden, in der Palaeontologie mit einem besonderen, für ihre praktischen Möglichkeiten "brauchbaren" Artkonzept zu operieren. Wir unterstreichen mit allem Nachdruck: So wie jede rezente Tier- und Pflanzenart heute in einzelnen Individuen existiert, so war auch in der Vergangenheit jedes fossilierte Lebewesen der Angehörige einer bestimmten evolutionären Art, die einmal als eine reale Einheit der Natur existiert hat.
Wir stehen damit vor der paradoxen Situation, einen Artbegriff für ein Fachgebiet zu fordern, dem seine genaue Anwendung prinzipiell verschlossen ist. Kommen wir in diesem Zusammenhang noch einmal auf das nach Willmann zitierte Beispiel des Brachiopoden Composita subtilita zurück (vgl. p. 295). Nach Willmann handelt es sich bei den verschiedenen Formen vom Unterkarbon bis in die Leonard-Stufe um Angehörige einer einzigen Art. Schon im Text zur Abbildung haben wir die Frage erhoben, wie es sich denn mit den Formen der letzten, der Guadelupe-Stufe, verhält. Hier rechnet der Verfasser offensichtlich mit der Möglichkeit einer Artaufspaltung. Ich möchte jedoch mit dem Maßstab heutiger Formenmannigfaltigkeit innerhalb vieler Arten noch einmal darauf hinweisen, dass keine Schwierigkeiten bestehen, die restlichen Formen ebenfalls zur selben Art zu zählen (vgl. p. 107f. u. 196f.). Bei vielen als Artaufspaltungen gedeuteten paläontologischen Befunden dürfte es sich um nichts weiter als unterschiedliche Ausschnitte des Variabilitätsspektrums der Arten handeln.
Wie zitiert, weist Ax darauf hin, dass man selbstverständlich 'eine hohe Übereinstimmung oder Identität in allen fossilisationsfähigen Strukturen als Indizien für die Zugehörigkeit verschiedener Individuen zu einer Art bewerten wird'. Das trifft auf die von Populationsgenetikern inzwischen auf Hunderttausende bezifferten Zwillingsarten jedoch nicht zu (viele davon sind durch absolute reproduktive Barrieren voneinander getrennt). "Ebenso selbstverständlich wird man signifikante Unterschiede im Merkmalsmuster verschiedener Fossilien als Ausdruck einer einstmaligen reproduktiven Isolation deuten und die entsprechenden Individuen als Vertreter verschiedener evolutionärer Arten hypothetisieren" - fährt Ax in seinem Kommentar fort. Wir wissen jedoch, dass eine ganze Palette signifikant unterschiedlicher Merkmalsmuster allein auf Rekombination von Genen (3. Mendelsche Regel) innerhalb der Art zurückgehen kann (vgl. p. 80), und wir müssen hier die Befürchtung äußern, dass eine solche Praxis doch wieder voll in den morphologischen Artbegriff zurückfallen könnte. Willmann geht jedoch hier einen Schritt weiter, wenn er den Vorschlag McAlesters (1962) zurückweist, aufgrund morphologischer Merkmale bei direkt von einem gemeinsamen Vorfahren abstammende Populationen auf die Entstehung reproduktiver Isolation zu schließen. "Die morphologische Divergenz spiegelt nicht direkt die genetische Verschiedenheit wieder: wir können nur bei sympatrischem Vorkommen die tatsächlichen Fortpflanzungsbeziehungen ermitteln" (Willmann, p. 153).
Aber auch bei der Sympatrie-Frage dürfte mit paläontologischem Material die Frage oft schwer zu entscheiden sein, ob eine absolute Reproduktionsbarriere vorliegt oder nicht. Formen wie die oben (pp. 78, 79, 170) ausführlich behandelten Geum-Arten Geum rivale und G. urbanum würden sich in der Regel in allochthonem und autochthonem Fossilmaterial wie zwei durch eine absolute Fortpflanzungsisolation voneinander getrennte Arten darstellen. Wie oben im Detail geschildert, sind die Forrren jedoch voll fertil. Denken wir weiter an das Kapitel der Kreuzungspolygone (pp. 170-195), insbesondere an die Gattungen Madia, Gilia und Elymus, so wird uns klar, dass die so unterschiedlichen Fertilitätsverhältnisse - von guter Fertilität bis zu vollständiger Sterilität zwischen den 'Arten' (ohne morphologische Korrelation) - paläontologisch nicht erfasst werden kann. Wie wir gesehen haben, ist das selbst bei rezentem Material oft nicht einfach, da verschiedene Linien ein- und derselben (morphologischen) Art völlig unterschiedliche Fertilitätsverhältnisse mit Linien einer anderen Art aufweisen können (vgl. oben p. 126, 130 die Verhältnisse bei Drosophila und p. 188f bei Mimulus sowie die auf der Seite 172 zitierten Ausführungen von Briggs und Walters (1984) und die daran anschließenden Kommentare). Auch die von Grant dokumentierten Fertilitätsunterschiede zwischen den verschiedenen Lebensformen der Pflanzen gehören hierher (vgl. pp. 174/175). Denken wir weiter an die zitierten Unterschiede zwischen der Frühlings- und Spätsommerform von Araschnia levana und anderen von der Jahreszeit abhängigen Schmetterlingsformen innerhalb der Arten sowie an den Geschlechtsdimorphismus im Tierreich allgemein (vgl. z.B. p. 157).
Willmann anerkennt diese Schwierigkeiten und unterstreicht sie mit einem Haeckelzitat über Fehleinordnungen von 'Jugendformen, Larven und Ammen', die als eigene Spezies, ja sogar 'als Glieder weitentfernter Familien oder selbst Klassen beschrieben' worden sind. Weitere Schwierigkeiten gibt es bei polymorphen Arten und der geographischen Variabilität (vgl. Willmann, pp. 106-109).
Bei morphologisch stark unterschiedlichen Drosophila-Arten von Hawaii finden wir "considerable interspecific fertility" (Carson), während zwischen den morphologisch kaum noch oder gar nicht unterscheidbaren Zwillingsarten derselben Gattung absolute Sterilität bestehen kann. Wie will man das paläontologisch unterscheiden? Kornfield et al. beschreiben die ökotrophisch und morphologisch stark differenzierten sympatrischen Cichliden in Cuatro Ciénegas. "By any phenetic criteria, the extreme distinctness of sympatric cichlids in Cuarto Ciénegas suggests the presence of two discrete species." (Vgl. p. 186 ). Welcher Paläontologe würde - auch bei Anwendung des Biospezies-Begriffs - bei solch starken anatomischen und ökotrophischen Unterschieden noch auf Angehörige einer Art schließen? Willmann meint jedoch zum Thema der Erfassbarkeit fossiler Biospezies im Zeitquerschnitt 1985, pp. 106/107: "Sympatrisches und synchrones Vorkommen erlauben die Schlußfolgerung, daß zwischen merkmalsverschiedenen Individuengruppen reproduktive Isolation besteht bzw. bestand." - Was bei den Cichliden-, Drosophila- (und tausend weiteren) Beispielen mit Sicherheit zu Fehlschlüssen führen würde. (Der Vollständigkeit halber sei jedoch vermerkt, dass der Autor im Anhang seines Buches (pp. 181/182) auch Beispiele zur diskontinuierlichen Variation innerhalb von Arten aufführt, was seine obige Aussage relativiert.)
Wiederholt haben wir jetzt schon die Zwillingsarten angesprochen. Da bei der undifferenzierten Verabsolutierung der Sterilitätsbarriere als Artabgrenzungskriterium Hunderttausende von Zwillingsarten akzeptiert werden müssen, handelt es sich hier für den Biospeziesbegriff (genauso wie für den evolutionären Artbegriff) in der Paläontologie um ein grundlegendes Problem, das Willmann ebenfalls anerkennt, wenn er p. 165 schreibt:
Wenn eine phylogenetische Aufspaltung, d.h. eine Speziation, übersehen wird, dann stimmt ein Taxon, das wir als Art bezeichnen, nicht mit dem Rahmen der natürlichen Biospezies überein. Es wird mehrere Biospezies bzw. Teile mehrerer Biospezies umfassen. Bei Zwillingsarten beispielsweise ist eine phylogenetische Aufspaltung ohne erkennbaren morphologischen Wandel erfolgt. Ein derartiges Speziationsereignis wird uns in der Paläontologie stets verborgen bleiben, und zwangsläufig fassen wir in einem solchen Fall zwei Biospezies zu einer taxonomischen Art zusammen.
Der Verfasser erkennt die Zwillingsarten ausdrücklich als eigene Arten an, wie er auch auf den Seiten 53/54 und 115 seiner Arbeit hervorhebt. Wenn nun heutzutage Hunderttausende von Zwillingsarten existieren (nach Schätzungen mancher Populationsgenetiker sogar Millionen, - vgl. White 1978), dann heißt das nach den Voraussetzungen der Synthetischen Evolutionstheorie, dass in der ganzen paläontologischen Zeitspanne Millionen, vielleicht sogar Milliarden von Zwillingsarten existiert hätten, die jedoch paläontologisch allesamt prinzipiell nicht erfassbar wären!
(Manchmal wird bei diesem Thema darauf hingewiesen, dass bei genauem anatomischen Studium von Zwillingsarten doch noch gewisse Unterschiede erkennbar werden - Mayr 1967, Ax 1984. Ein solcher Einwand würde jedoch für die vorliegende Aussage keine Rolle spielen, weil auch die mit viel Mühe nachgewiesenen und wiederidentifizierten Unterschiede so gering bleiben, dass sie von der Mendelschen Rekombination innerhalb der meisten Arten um ein Vielfaches übertroffen werden.)
Auf der Seite 85 seiner Arbeit versucht Willmann im Anschluss an einen Einwand Cronquists (vgl. p. 303) seinen Biospeziesbegriff für die Praxis zu rechtfertigen, indem er schreibt:
Daß sich aus einem theoretisch einwandfrei definierten Begriff in der Praxis Schwierigkeiten ergeben können, hat seit jeher zu Kontroversen geführt. Zu leicht wird vergessen, daß uns der Begriff in seiner theoretisch korrekten Begründung das Ziel zeigt, das es anzustreben gilt. Wie Hennig (1969: 36) in einem anderen Zusammenhang ausführte, kann niemand vorhersagen, ob in der Praxis auftretende Schwierigkeiten, die uns heute noch den Weg zu diesem Ziel verbanen, nicht doch eines Tages überwunden werden können. Wenn wir aber von vornherein einem praxisorientierten Artbegriff folgen würden, dann bestünde kein Anreiz, nach Methoden zu suchen, die zur Überwindung solcher Schwierigkeiten führen könnten.
- Es erscheint mir jedoch sehr fraglich, ob mit dem Biospeziesbegriff ein Anreiz besteht, nach Methoden zur Überwindung der in der Paläontologie bestehenden Schwierigkeiten zum Thema Zwillingsarten zu suchen.
Die reproduktive Isolation macht Willmann zur einzigen und absoluten Grundlage seines gesamten Artkonzepts. Er hebt an verschiedenen Stellen seines Buches hervor, dass er damit die "absolute Isolation" meint und geht damit über den Neodarwinismus hinaus (vgl. das Möwenbeispiel p. 93). Er hebt seine Übereinstimmung mit Klausnitzer und Richter hervor, die 1979, p. 239 schrieben: "Eine fertile Nachkommen ergebende Artbastardierung ist unmöglich." Schwache Fortpflanzungsisolation lehnt er ab, denn bei solcher "gibt es Übergangspopulationen und damit keine natürliche Grenze" (p. 47). "Wenn wir also als Art das bezeichnen wollen, was in der Natur als kleinste, von allen anderen Populationen biologisch isolierte Einheit besteht, und wenn sich das Artkonzept auf reale Objekte beziehen soll, wie es in meinen Augen für einen naturwissenschaftlichen Forschungszweig unabdingbar ist, dann muß mit "reproduktiver Isolation" die absolute Isolation gemeint sein" (ebenfalls p. 47).
Sehen wir uns näher an, was der Autor mit der absoluten Isolation meint. So ist es z.B. noch möglich, dass zwei Biospezies unfruchtbare Hybriden hervorbringen, und im Falle der Allopolyploidie erzeugen zwei Spezies eine dritte (p. 47). Bei der bereits zitierten Besprechung des Syngameonkonzepts (vgl. p. 204 der vorliegenden Arbeit) hat der Autor noch konsequent sein Biospezieskonzept vertreten: Zusammenfassung aller in einem Syngameon vertretenen 'Arten' zu einer einzigen Art. Beim Einwand Cronquists, 'daß Dobzhanskys Formulierung des Artbegriffs für die Botanik wenig wert habe', da sie bedeuten würde, 'daß eine Vielzahl von Arten und sogar Gattungen z.B. der Orchideen zu einer Art zusammenzufassen wären' (der Schutz gegen Hybridisierung beruht hier weitgehend auf dem Zusammenspiel mit bestimmten Bestäubern, und man kann durch Kreuzung Hybriden zwischen sehr verschiedenen Arten erzeugen), schreibt Willmann (p. 85): "Cronquist geht nicht darauf ein, daß die Anpassung an bestimmte Bestäuber als ein vor der Befruchtung eingreifender Isolationsmechanismus anzusehen ist." Die Arten sind nach Willmann also nicht allein durch Sterilitätsschranken isoliert, und er schreibt für die Botanik: "Daß ähnlich wie im Tierreich auch ganz andere Isolationsmechanismen existieren, wurde erst sehr spät erkannt" (ebenfalls p. 85).
Zur "absoluten Isolation" gehören also auch die präzygotischen Isolationsmechanismen, von denen wir oben in aller Ausführlichkeit klargelegt haben, dass eine solche Isolation in der Regel keineswegs absolut ist (vgl. pp. 69-120). Gerade bei den Orchideen gibt es auch in der Natur zahlreiche Hybridpopulationen - vgl. oben die Beispiele bei der Gattung Ophrys pp. 107-112 (siehe auch den Bartfaden p. 106).
Was bei den Orchideen generell möglich ist, soll die folgende Liste aus Encke, Buchheim und Seybold (1980, pp. 62 - 64) verdeutlichen, zu der vorauszuschicken ist, dass sämtliche Gattungen, vor denen ein Multiplikationszeichen steht, Hybridgattungen (Gattungsbastarde) sind:
Buttler schreibt 1986, p. 260:
Wie keine andere Familie der Gefäßpflanzen neigen Orchideen zur Bastardbildung. Kreuzungsbarrieren bestehen innerhalb der Gattungen kaum, auch nicht zwischen Gattungen, wenn sie zur selben Tribus gehören. ...Wo immer Arten derselben Gattung oder Tribus eng benachbart wachsen und sich nicht durch die Blütezeit trennen, kann man mit Bastarden rechnen.
Über die Hälfte aller bei den Orchideen heutzutage gezählten Gattungen sind Hybridgattungen (179 von 351) [Es scheint in der Systematik kaum feste Zahlen zu geben: Bei Willis (1985) habe ich 287 Hybridgattungen gezählt, dafür wartet er jedoch mit insgesamt 735 Gattungen bei den Orchideen auf. Das hieße dann rund 40% Hybridgattungen.], d.h. sie sind aus Kreuzungen zwischen anderen Gattungen hervorgegangen. Eine spezielle Studie wäre zur Beantwortung der Frage nötig, wieviele der übrigen, nicht durch Kreuzung entstandenen Gattungen miteinander kreuzbar und wieviele Hybriden davon fertil und wieviele steril sind. So wie die rund 450 Hunderassen samt Wolf, Schakal und Coyote (mit allen ihren Unterarten) genetisch eine einzige Art bilden, so dürfte sich die Vielfalt der über 20 000 morphologischen Arten der Orchiden auf wenige Gattungen und Arten reduzieren.
Zahlreiche Bastardierungen sind bei den Orchideen auch in der Natur beobachtet worden, so dass von einer "absoluten Isolation" weder in der Natur noch in der züchterischen Praxis die Rede sein kann. Von Willmann wird hier also ein keineswegs absoluter Isolationsmechanismus als artentrennend anerkannt. Schon auf den Seiten 43/44 seines Werkes klingt dieser Punkt an, wenn er schreibt: "Bei absolut wirksamen, vor der Paarung eingreifenden Isolationsmechanismen bringen die potentiellen Gechlechtspartner zweier Arten keinerlei Nachkommen hervor." Das auf den Seiten 88-93 abgehandelte Larus-Beispiel wird nicht etwa wegen der Fertilitätsfrage bei den Hybriden der verschiedenen Möwenformen als 'noch nicht vollendete Aufspaltung' eingeordnet, sondern weil eine ununterbrochene Fortpflanzungsgemeinschaft über die zwischengeschalteten Populationen' gegeben ist. "Solange aber die Aufspaltung nicht abgeschlossen ist, solange existiert auch keine absolute Fortpflanzungsisolation" (p. 48). Wann wäre denn nun die Aufspaltung abgeschlossen? Nach den vorliegenden Ausführungen von Willmann logischerweise, wenn die 'ununterbrochene Fortpflanzungsgemeinschaft über die zwischengeschalteten Populationen' unterbrochen wird, wenn z.B. durch Veränderung der Ökosysteme eine dieser Populationen ausfallen ("ausgeschaltet") würde. Genetisch hätte sich zwischen den Endpopulationen deswegen zwar nichts verändert, sie würden jetzt aber zu "absolut" voneinander getrennten Arten gehören. Und an diesem Punkte setzen nun all unsere Einwände wieder ein, die wir gegen eine solche Auffassung geltend gemacht haben (vgl. pp. 75, 77, 94), und die wir an dieser Stelle nicht noch einmal wiederholen möchten. Beim Larus-Beispiel kommt noch hinzu, "daß vollständige Kontinuität der Populationen nicht gegeben ist. Es sind häufige Lücken in der Reihe der Bevölkerungen zwischen Westeuropa und Ostasien da, und einige von ihnen sind soweit ausgeprägt, daß die Autoren [Stresemann und Timophéeff] sich veranlaßt sahen, den Larus-argentatus-Komplex in drei Arten aufzugliedern" (vgl. Mayr, p. 90 oben). (Da dieser Punkt von so vielen Autoren übersehen wird, schien es mir sinnvoll, diese Ausführungen Mayrs hier zu wiederholen.) Nach den Willmannschen Voraussetzungen handelt es sich bei diesem Beispiel schließlich doch noch um (mindestens) zwei verschiedene Arten.
Oben (p. 296) hatten wir Willmann (und anschließend Ax) mit der Auffassung zitiert, dass 'zwei allochrone Populationen derselben evolutiven Linie auch dann in ein und dieselbe Spezies gestellt werden müssen', "wenn z.B. aus Gründen der Konstruktion ausgeschlossen werden kann, daß ihre Vertreter sich erfolgreich verpaaren konnten..." Der Autor vergleicht dann diese Situation mit den "ring-species", wo 'sogar Populationen eine Biospezies bilden, die nachweislich reproduktiv voneinander isoliert sind'. Da die oben zitierte Aussage Mayrs für die Rassenkreise generell gilt, kann Willmann nach seinen eigenen bisherigen Voraussetzungen diesen Vergleich zur Klärung der paläontologischen Situation nicht mehr weiter einsetzen, weil es sich durch die Diskontinuität der Populationsketten bei den Rassenkreisen dann doch um zwei oder mehrere Arten handelt - es sei denn, der Autor wäre bereit, aufgrund der zahlreichen oben aufgeführten genetisch-systematischen Probleme die präzygotischen Isolationsmechanismen als artentrennend fallenzulassen und den Artbegriff weiterzufassen.
Dasselbe gilt für die ganze Problematik der Zwillingsarten. Wie wir oben detailliert ausgeführt haben, handelt es sich bei diesen um Arten 'sekundären Ursprungs', wobei mit der Aufrechterhaltung des Artstatus eine ganze Serie von Problemen und verworrenen Konsequenzen verbunden ist, die nur aus der Welt geschafft werden kann, wenn man solche Linien unter den Artbegriff subsumiert. Mit der auf den Seiten 210-269 diskutierten Differenzierung in primäre und sekundäre Arten und der Subsumption aller durch Mendelsche Spaltung und Rekombination bedingten präzygotischen Isolationsmechanismen sowie sämtlicher durch Degeneration verursachte (=sekundäre) Sterilitätsbarrieren unter den Artbegriff, entfällt hingegen diese Problematik sowohl für die Neontologie als auch die Paläontologie.
Übrigens hat Willmann die auf den Seiten 113-115 zitierten und diskutierten Einwände gegen den neodarwinistischen Artbegriff von Ehrlich und Raven nicht widerlegt, wenn er in seinem Buch kommentiert (p. 50):
Die genannten Autoren gingen von der Überlegung aus, daß der Genfluß, der Austausch von Genmaterial zwischen benachbarten Populationen einer Art, eine wesentliche geringere Rolle spiele als oft angenommen wird. So komme geographisch separierten Populationen oft eine Bedeutung zu, die der von Arten gleiche. Aber das steht weder im Gegensatz zum Biospezies-Konzept, noch rührt es an seiner Gültigkeit oder an der evolutiven Bedeutung der Arten (vgl. Mayr 1969b: 318). Zweifellos sind die Teilpopulationen einer Art (oft = Unterart) für die Evolution besonders wichtig: Eine Biospezies, erst einmal entstanden, tendiert mit ihrer Ausbreitung dazu, in Teilpopulationen und letztlich in neue Arten zu zerfallen. Aber damit wird nichts anderes als der Kreis Biospezies - infraspezifische Aufgliederung - Aufspaltung einer Biospezies in Tochterarten (= neue Biospezies) geschlossen...
Wie schwierig die Situation für den neodarwinistischen Artbegriff wird, wenn die ursprünglich als Kontinuum gedachten Ringspezies in der Realität aus diskontinuierlichen Populationen bestehen, haben wir ja gerade für Willmanns Biospezies-Konzept aufgezeigt (s.o.): Er erkennt den Artstatus der Endglieder beim Larus-Beispiel nicht an, weil (wie er meint) eine 'ununterbrochene Fortpflanzungsgemeinschaft über die zwischengeschalteten Populationen' besteht und daher die Aufspaltung nicht abgeschlossen sei. Solange aber die Aufspaltung nicht abgeschlossen ist, solange existiert auch keine absolute Fortpftanzungsisolation. Und solange dies nicht der Fall ist, besteht die Gesamtheit der Populationen eines solchen Formenkreises als einheitliche Biospezies" (pp. 48/50). - Worauf der Hinweis kommt, dass Larus argentatus und Larus fuscus lediglich Subspezies ein und der selben Art sind.
In dem Augenblick also, indem die Endglieder nicht mehr durch eine ununterbrochene Fortpflanzungsgemeinschaft miteinander verbunden sind, hätten wir nach diesem Artbegriff wenigstens zwei Spezies, nämlich die Endglieder dieses Ringes.
Der entscheidende Punkt bei den Einwänden von Ehrlich und Raven und vieler anderer neuerer Autoren (vgl. pp. 114-116) ist die Tatsache, dass ein großer Teil der als Kontinuum gedachten Speziesverbreitung in der Realität aus zahlreichen, oft Hunderten und Tausenden kleiner Populationen besteht, zwischen denen es wenig oder gar keinen Genfluss gibt, - und dieses Phänomen ist für eine große Zahl von Beispielen gefunden worden, ohne dass sich irgendwelche prä- und/oder postzygotische Isolationsmechanismen herausgebildet hätten. Wichtig ist dabei noch festzuhalten, dass das sowohl für große Entfernungen als auch für große Zeiträume gilt (vgl. p. 115).
Nach den Vorstellungen der Synthetischen Evolutionstheorie hingegen wird mit der Unterbrechung des Genflusses auch die Entstehung prä- und/oder postzygotischer Isolationsmechanismen mit häufig darauf folgender morphologischer Differenzierung impliziert, und zwar nach der Regel: Einschränkung des Genflusses -> Bildung prä- und postzygotischer Isolationsmechanismen -> weitere Einschränkung des Genflusses -> weitere Differenzierung (einschließlich morphologisch-anatomischer Merkmale) -> verstärkte bis fast vollständige Unterbrechung des Genflusses -> verstärkte Differenzierung auf allen Ebenen -> absolute Fortpflanzungsisolation -> Bildung verschiedener Arten (und darauf folgend in größeren Zeiträumen), Gattungen, Familien, Ordnungen etc..
Statt dessen finden wir häufig eine hundert- und tausendfache Einschränkung des Genflusses innerhalb der Art durch diskontinuierliche geographische Verbreitung in kleineren und größeren Populationen, und dennoch keine Differenzierung zwischen den Populationen. (Auf der anderen Seite gibt es wieder starke morphologische Unterschiede innerhalb und zwischen Populationen ein- und derselben Art, aber keine prä- und postzygotischen Isolationsmechanismen (vgl. pp. 186/187), und die Zwillingsarten schließlich können reproduktiv absolut voneinander isoliert sein, ohne irgendwelche morphologischen Differenzen aufzuweisen.)
Die neodarwinistische Idee, dass in der Regel die Differenzierungsprozesse an die Frage des Genflusses und dessen schrittweise Unterbrechung gekoppelt seien, ja dass die ganze Artbildungsfrage und implizit der Ursprung aller Lebensformen in großen Zeiträumen, von der Genfluss-Regulation abhängt, steht im Gegensatz zu zahlreichen pflanzen- und tiergeographischen, morphologischen und genetischen Tatsachen (s.o.). (Wollte man die oft Tausende geographisch voneinander solierten Populationen einer Spezies als werdende Arten betrachten, so käme man bei Betrachtung der gesamten Orgarismenwelt auf Milliarden von Spezies in statu nascendi).
Fazit: Die von Ehrlich und Raven (und vielen weiteren Autoren - vgl. p. 115) aufgeführten pflanzen- und tiergeographischen Gegebenheiten stehen sehr wohl im Gegensatz zum Biospezies-Konzept und rühren an seiner (allgemeinen) Gültigkeit und der evolutiven Bedeutung dieses Artbegriffs.
Oben hatte ich Willmanns Behauptung zitiert, dass eine Art vor allem in Relation zu ihrem nächstverwandten Taxon eine Art ist. Nach der 'Artaufspaltung' sind die beiden Populationen als neue Arten "und die Stammart als erloschen aufzufassen". Dieser letztere Punkt wird von kladistischen Autoren auch dann noch aufrechterhalten, wenn die Stammart unverändert weiterexistiert (vgl. auch Ax 1984; ursprünglich hatte auch Hennig diesen Gedanken vertreten, vgl. Willmann 1985, p. 170).
Die beiden Artbildungsmodi im Sinne des Kladismus hat Willmann folgendermaßen veranschaulicht (p. 158):
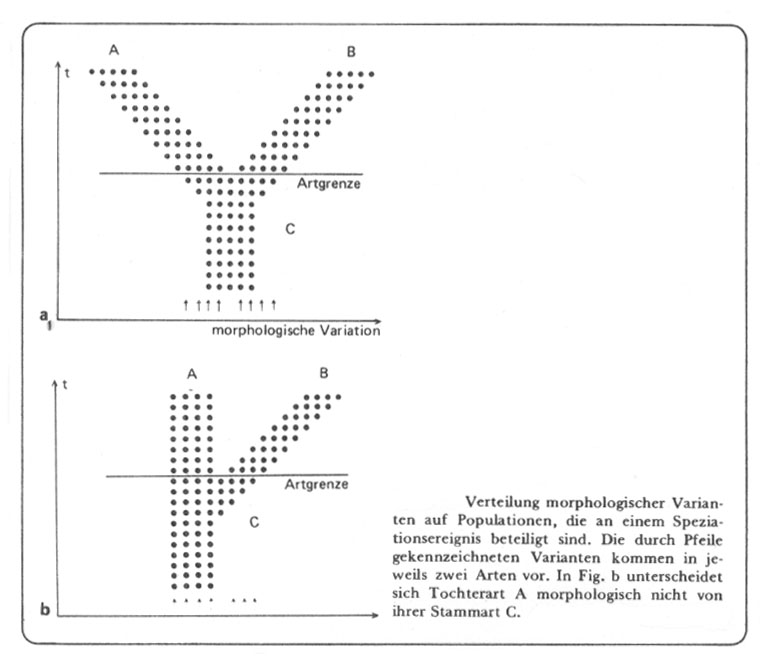
Abb. 46: Aus Willmann 1985, verkleinert.
Genetisch sieht die Situation in der Regel ganz anders aus: Die Art C enthält in beiden Fällen ein umfangreiches Rekombinationspotential (bei nur 30 heterozygoten Genpaaren mindestens 1 073 741 824 phänotypisch verschiedene homozygote 'Varianten' - vgl. p. 80). Unter den gegebenen geographisch-ökologischen Bedingungen kann sich davon nur ein Teil mit größerer Individuenzahl behaupten, und paläontologisch seien (schematisiert) nur 4 bis 5 Gruppen morphologisch gut unterscheidbarer Varianten nachweisbar (obige Abb.).
Durch Verlagerung ökologischer Gegebenheiten und Besiedlung neuer Lebensräume kommt es zu einer Divergenz der mit größeren Individuenzahlen vertretenen Rekombinanten in den verschiedenen Gebieten (a). In der Abbildung b bleiben die ökologischen Bedingungen für die Art C so konstant, dass keine Verlagerung in der Rekombinantenhäufigkeit in der Zeit t auftritt, sondern nur bei Besiedlung neuer Lebensräume. In beiden Fällen hat sich am Rekombinationspotential noch nichts geändert, - es werden nur unterschiedliche Ausschnitte aus diesem Potential unter den verschiedenen Umständen verwirklicht (vgl. oben pp. 78/79 das Geum-Beispiel). Von Artbildung kann dabei nicht die Rede sein, auch wenn bei Pflanzen die Populationen unterschiedlicher Rekombinanten von unterschiedlichen bestäubenden Insekten besucht werden (wie bei
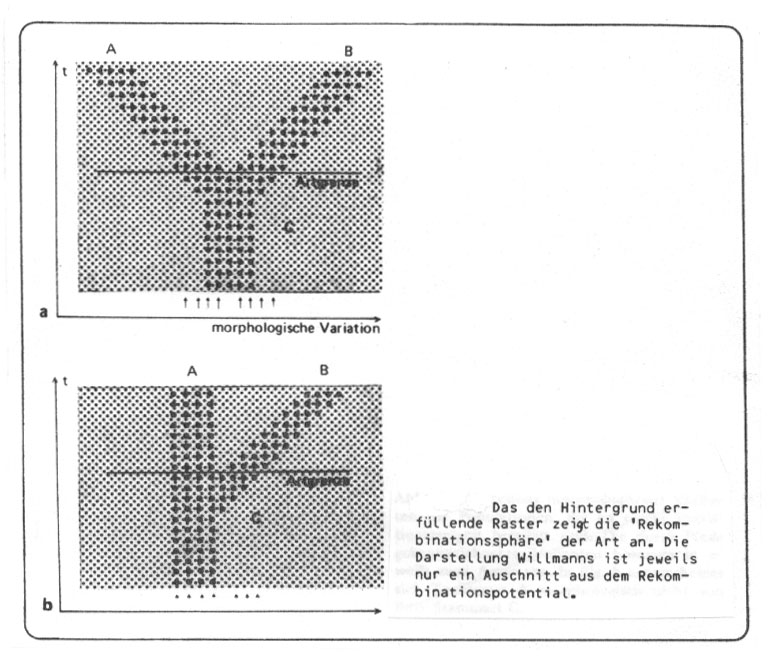
Abb. 47: Aus Willmann 1985, verändert.
den Orchideen) oder Prägung auf die Eltemrekombinante stattfindet (wie bei vielen Vögeln) etc. (vgl. die Diskussion der prä- und postzygotischen Isolationsmechanismen).
Immerhin werden Verlagerungen aus dem Rekombinantenpotential ohne "Artaufspaltung" von Ax, Willmann u.a. nicht als Artbildungsprozess aufgefasst. In der folgenden Abbildung wird zunächst die häufig von Paläontologen gegebene Deutung der Artentransformation wiedergegeben und dann die Anwendung des Biospezies-Konzepts klargestellt:
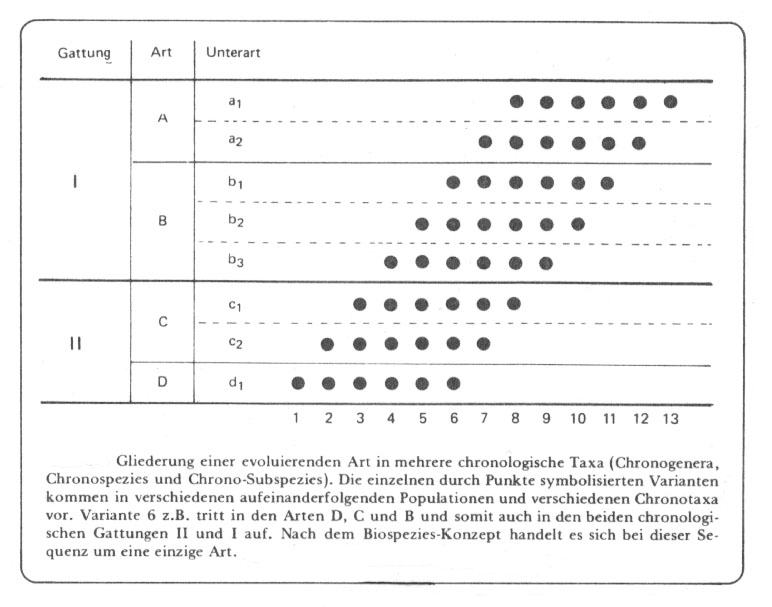
Abb. 48: Aus Willmann 1985, verkleinert.
Dass es sich bei solchen Beispielen nach dem Biospezies-Konzept um eine einzige Art handelt ist schon ein großer Fortschritt gegenüber dem morphologischen Artbegriff in der Paläontologie, der nach diesem Schema immerhin noch 4 Arten und 2 Gattungen unterschieden hätte. Nicht verständlich ist mir jedoch die Formulierung von "einer evoluierenden Art". Wie schon erwähnt, wird in solchen Fällen häufig auch von Artumwandlung, phyletischer Evolution, phyletischer Speziation und Transformation von Arten in der Zeit gesprochen - die Deutung der Transformation wie auch der Begriff der "evoluierenden Art" ist häufig nichts als die Verkennung des "ungeheuren Umfang" des genetischen Potentials, von dem unter den gegebenen geographisch-ökologischen Bedingungen immer nur ein geringer Teil für eine größere Individuenzahl einer Population realisiert ist. Die Kenntlichmachung dieses Potentials in der nachstehenden Abbildung (Kreise) möchte diesen Gedanken veranschaulichen:
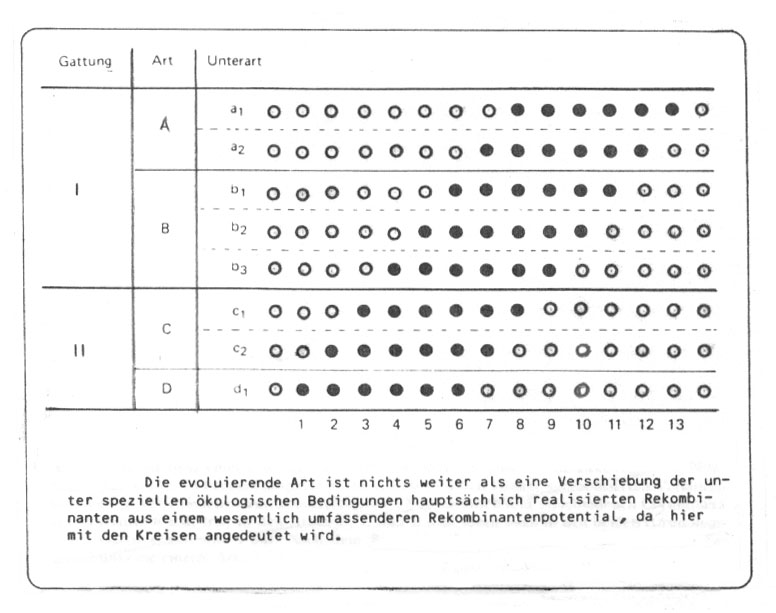
Abb. 49: Aus Willmann 1985, verändert.
In der genetischen Praxis erheben sich übrigens mit dem Artabgrenzungsversuch durch 'Aufspaltung' der Arten einige Fragen, die wir im folgenden kurz umreißen möchten: Eine Stammlinie (Ausgangslinie) wird mit mutagenen Agenzien behandelt und durch Abbau und Umbau genetischer Strukturen liefert sie schließlich zahlreiche neue Linien, von denen einige mit der Ausgangslinie nur noch schwer kreuzbar sind, so dass sie in der Natur 'absolut' voneinander isoliert wären. In diesem Falle müssten nach Meinung vieler Kladisten nicht nur diese letzteren (neuen) Linien neue Artnamen, sondern auch die immer noch vorhandene - (und genetisch vor und nach diesem Ereignis bzw. Ereignissen völlig identische) Ausgangslinie einen neuen Artnamen erhalten.
Wenn z.B. die Translokationslinien der Kulturvarietät 'Dippes gelbe Viktoria' der Erbse (Pisum sativum) nach weiterer Behandlung mit mutagenen Agenzien schließlich von der Ausgangsform 'absolut' isoliert sind, dann sind nicht nur diese neuen Linien, sondern auch die Ausgangslinie mit einem Mal neue Arten, die dann entsprechend auch einen neuen Artnamen erhalten müssten.
Die Kulturvarietät 'Dippes gelbe Viktoria' gehörte dann plötzlich nicht mehr zu Pisum sativum, sondern (sagen wir) zu Pisum viktorianum. Und dasselbe träfe dann auf sämtliche andere Kulturvarietäten der Erbse zu: sie alle gehörten plötzlich zu dieser neuen Art - , aber nur solange, bis eine weitere 'Artaufspaltung' stattfände. Schließlich hätte ein und dieselbe genetisch reine Linie ein Dutzend und noch mehr verschiedene Artnamen, die alle jeweils nur für einen bestimmten Zeitraum gelten würden, - was dann etwa folgendermaßen aussehen könnte: die Translokationslinie 405 b aus 'Dippes gelbe Viktoria', jetzt Pisum cladicum, vorher - vom 3.8.81 bis 11.12.85 - P. multinomicum, vom 18.7.73 - 3.8.81 P. molestum, vom 1.8.71 12 Uhr 30 bis 18.7.73 16 Uhr 48 P. elongellum, vom 12.11.61 bis 1.8.71 P. enduratum usw. usf. - mit dem die Aufzählung abschließenden Hinweis "von Linné im 18. Jahrhundert ursprünglich als Pisum sativum bezeichnet" - erhält jetzt den Artnamen P. degeneratum. Dieser Artname gilt bis zur nächsten Artaufspaltung, die allerdings bereits vor einer Woche (Müller mündliche Mitteilung) induziert wurde. Namenrevisionen werden nachgeliefert. Die Folge wäre letztlich die dauernde Umbenennung aller möglichen Spezies, die jedoch genetisch zum großen Teil unverändert weiterexistieren würden. Kein vernünftiger Biologe könnte ein solches nomenklatorisches Chaos verantworten, aus dem sich selbst Spezialisten nach einer gewissen Zeit dauernder Umbenennung kaum noch zurecht finden würden.
Wie wir oben jedoch schon ausführlich gezeigt haben (pp. 121-168), sind solche Fälle von Chromosomenumbau und genetischem Strukturabbau unter den Artbegriff zu subsumieren, und die soeben aufgeführten weiteren Schwierigkeiten dürften die Richtigkeit dieser Schlussfolgerung noch bestärken.
Willmann hält die Ablehnung, nach "Artaufspaltung" unveränderte Linien mit neuen Artnamen zu versehen, für typologisches Denken. Eine reine Linie ist aber nicht typologisch, sondern genetisch durch die Homozygotie definiert.
Wiederholt haben wir die Kritik von Willmann und Ax zur Auffassung zitiert, dass allein durch Transformation von Arten in der Zeit ("Artumwandlung, phyletische Evolution, phyletische Speziation") keine neuen Arten entstehen könnten, da nach ihrer Meinung ein "Spaltungsprozeß" zur Artabgrenzung gehört.
Mit der Bereitschaft, mehrere kontinuierlich ineinander übergehende Taxa (im obigen Schema Arten und Gattungen) zu einer Art zusammenzufassen, bietet der "evolutionäre Artbegriff" trotz seiner fragwürdigen Artabgrenzungsdefinition einen interessanten Ansatz zur Objektivierbarkeit des paläontologischen Artkonzepts. Wie hervorgehoben, spielt es bei "nicht aufspaltenden" Arten keine Rolle, wie weit sich die Endpopulation (wie die Unterart a1 in Abb. 49) von der Ausgangspopulation (Unterart d1 derselben Abbildung) morphologisch-anatomisch und ethologisch entfernt hat. Die zur Artabgrenzung herangezogenen prä- und postzygotischen Isolationsmechanismen gelten nur im 'Zeitquerschnitt'. Im Längsschnitt hingegen können sich die Populationen so stark von einander unterscheiden, dass nach allen vorliegenden Parametern (Unterschiede in der Körpergröße, dem Bau der Geschlechtsorgane, in der Blütenform etc.) vollständige Isolation zwischen ihnen bestehen würde, wenn man sie auf die gleiche Zeitebene transponieren könnte. D.h. die gleichen Unterschiede, die als absolut artentrennend bei diskontinuierlicher Verteilung im geographischen Raum gelten, verlieren ihre Gültigkeit vollständig in der Zeit.
Das letztere ist jedoch konsequent gedacht, denn wir haben es in diesen Fällen mit einer Abfolge von genetisch direkt zusammenhängenden Populationen zu tun, und alle morphologischen Artabgrenzungen sind in solchen Fällen Willkür.
Eine Chance zur Annäherung an einen objektiven Artbegriff in der Paläontologie besteht nun darin, solche morphologisch-anatomischen Unterschiede und Abstände, die schließlich zwischen kontinuierlich zusammenhängenden Populationen auftreten, zu einem (ersten) Maßstab zur Beantwortung der Frage nach dem Umfang des Artbegriffs in der Zeit zu machen. Diese Abstände dürften jedoch von Ordnung zu Ordnung und Klasse zu Klasse unterschiedliche Ausmaße haben, so dass eine Extrapolation etwa über die Ordnungsgrenzen hinaus nicht ohne weiteres zu rechtfertigen wäre.
Es handelt sich in der paläontologischen Realität in der Regel um morphologische Arten eines Genus, die Kontinuität kann aber auch mehrere morphologisch definierte Genera umfassen.