
DIE FRAGE NACH DEN ANWENDUNGSMÖGLICHKEITEN DES LAMPRECHTSCHEN ARTBEGRIFFS IN DER SYSTEMATISCHEN PRAXIS
Folgende Einwände könnten von systematischer Seite erhoben werden:
Welche Bedeutung könnte einem Artbegriff beigemessen werden, der in der systematischen Praxis kaum anwendbar ist? Sollten denn jedesmal erst in jahrzehntelanger Arbeit die Erbgänge an Zehn- bis Hunderttausenden Pflanzen und Tieren untersucht werden, bevor die Frage entschieden werden könnte, ob es sich bei den Individuen zweier Linien um Angehörige derselben Art oder tatsächlich zweier verschiedener Arten handelt?
Selbst wenn wir nach genauesten Untersuchungen feststellen, dass bestimmte Merkmale nicht reziprok überführbar sind, könnte es sich nach der obigen Diskussion ja immer noch um sekundäre Arten durch Abbau genetischer Strukturen handeln (vgl. p. 237), die dann auch wieder unter der einen primären Art subsumiert werden müssten.
Darüber hinaus müssten zahlreiche Organismen, die in vielen Merkmalen und Merkmalskomplexen stark verschieden und somit klar unterscheidbar sind, zu einer Art zusammengefasst werden, was nun die tertiären Nomenklaturlisten fast uferlos erweitern würde.
Manche Linien sind durch genetische Abbauprozesse soweit degeneriert, dass sie sowieso nicht miteinander kreuzbar sind und deshalb die systematische Frage auf diesem Wege gar nicht entschieden werden kann (vgl. Beispiele pp. 131-135).
Ist es angesichts dieser Problematik nicht sinnvoller, den systematischen status quo beizubehalten und den genetisch-Lamprechtschen Artbegriff den Genetikern zu überlassen?
Ad 1. Durch die vor allem in unserem Jahrhundert durchgeführten, umfangreichen genetischen Untersuchungen mit nahverwandten 'Arten' liegt bereits ein großes Material vor, das sozusagen nur 'darauf wartet', terminologisch auf die genetische Ebene gebracht zu werden (ich möchte auch an die über 1000 Hybridpopulationen erinnern, die von Kerner schon 1891 aufgelistet wurden).
An je einem Beispiel aus dem Pflanzen- und Tierreich sei die Durchführung dieser Aufgabe veranschaulicht:
Oben haben wir das 'Syngameon' der Eichen nach Grant (1981) zitiert (vgl. p. 175). Nach herkömmlicher systematischer Einteilung unterscheiden wir hier folgende Arten (in alphabetischer Reihenfolge):
|
1. |
Quercus alba |
10. |
Quercus minima |
|
2. |
Quercus austrina |
11. |
Quercus muehlenbergii |
|
3. |
Quercus bicolor |
12. |
Quercus oglethorpensis |
|
4. |
Quercus chapmanni |
13. |
Quercus petraea |
|
5. |
Quercus durandii |
14. |
Quercus prinoides |
|
6. |
Quercus lyrata |
15. |
Quercus prinus |
|
7. |
Quercus macrocarpa |
16. |
Quercus robur |
|
8. |
Quercus margaretta |
17. |
Quercus stellata |
|
9. |
Quercus michauxii |
18. |
Quercus virginiana |
Aufgrund der oben besprochenen Daten reduzieren sich diese 18 Arten genetisch aller Wahrscheinlichkeit nach auf eine einzige Art (Q. chapmanni und oglethorpensis sind jedoch nach der Darstellung Grants noch nicht in das 'Syngameon' integriert und müssten demnach noch untersucht werden).
Um diese 18 Arten terminologisch auf die genetische Ebene zu bringen, gehen wir folgendermaßen vor: Nach den Nomenklaturregeln nehmen wir die zuerst beschriebene Art (in diesem Falle Quercus robur L.) und beschreiben die übrigen mit der tertiären Nomenklatur (wie das ja heute bei vielen Pflanzen- und Tiergruppen sowieso schon üblich ist):
|
1.a) |
Quercus robur robur |
j) |
Quercus robur michauxii |
|
b) |
Quercus robur alba |
k) |
Quercus robur minima |
|
c) |
Quercus robur austrina |
l) |
Quercus robur muehlenbergii |
|
d) |
Quercus robur bicolor |
m) |
Quercus robur oglethorpensis |
|
e) |
Quercus robur chapmanni (?) |
n) |
Quercus robur petraea |
|
f) |
Quercus robur durandii |
o) |
Quercus robur prinoides |
|
g) |
Quercus robur lyrata |
p) |
Quercus robur prinus |
|
h) |
Quercus robur macrocarpa |
q) |
Quercus robur stellata |
|
i) |
Quercus robur margaretta |
r) |
Quercus robur virginiana |
Erinnern wir uns an das treffende Wort Kaestners, dass 'Bildung das Bewußtsein der Zusammenhänge ist', so ist die botanisch-genetische Bildung mit der zweiten Aufführung unmittelbar gegeben: Die Art Quercus robur umfasst mindestens 16 von Systematikern als verschiedene Arten geführte Ökotypen, die alle miteinander fertile Nachkommen haben bzw. haben können (der europäische Teil des 'Syngameons' ist dabei noch gar nicht berücksichtigt). Die Ökotypen selbst sind von den Systematikern detailliert beschrieben worden. Die Namen der Autoren können weiterhin hinter den Namen der Ökotypen aufgeführt werden.
Die Ökotypen bzw. Subspezies werden jetzt also nicht mehr als eigene voneinander getrennte Arten aufgeführt, die sich vielleicht schon vor Jahrmillionen voneinander getrennt und nichts mehr miteinander zu tun hätten, sondern der realgenetische Zusammenhang wird mit der zweiten Aufführung unmittelbar vor Augen geführt: die vorher als artentrennend eingestuften Merkmale sind von jeder Art in alle anderen Arten und vice versa überführbar.
Als zoologisches Beispiel wollen wir die Bärenfamilie (Ursidae) nehmen:
Einleitend sei erwähnt, dass Merriam (1918) 78 Arten der nordamerikanischen Braun- und Grizzlybären unterschied (nach Willmann 1985).
Matthews schreibt über den Braunbären Ursus arctos (1972, p. 477):
Die Art ist in der Größe, der Färbung und auch in der Lebensweise sehr variabel.
...So vergab Merriam allein 86 Artnamen an nordamerikanische Braunbärformen, die er dort unterscheiden zu können glaubte, die aber heute zumeist nicht einmal mehr als Unterarten anerkannt werden.
Wir möchten uns mit der Klassifizierung der Bären folgender Autoren beschäftigen: Gray 1971 (I); Grzimek 1975/1979 (II); Stains 1974/1982 (III); Mohr 1972/1982 (IV); Wurmbach 1971 (V); Petzsch 1966/1974 (VI); Matthews 1971/1972 (VII); Manaranche 1984 (VIII).
Die Gattungen werden zunächst nach Gray aufgeführt; Übereinstimmungen, Synonyme und Abweichungen sind der Tabelle 4 zu entnehmen.
Anschließend haben wir die Bärenkreuzungen nach Gray aufgeführt (Tabelle 5). Das erste, was wir aus genetischer Sicht feststellen können, ist die Fragwürdigkeit der Klassifikation in die 6 Bärengattungen. Das wird auch von systematischer Seite zunehmend anerkannt: 3 der 8 Autoren führen den Eisbären schon nicht mehr als Thalarctos maritimus, sondern als Ursus maritimus. Und nur noch einer der Autoren schreibt Euarctos americanus statt (richtig) Ursus americanus.
Da die unter 1. und 2. aufgeführten Bärenformen der Tabelle 5 miteinander Nachkommen hervorbringen können, erscheint es vernünftig, sie zunächst einmal zur selben Gattung zu stellen:
Ursus malayanus
Ursus ursinus
Ursus thibetanus
(Streng genommen fehlt jedoch noch ein Glied in der Kette, nämlich eine Kreuzung zwischen Selenarctos und Melursus oder Helarctos. Wir gehen vorläufig von der Annahme aus, dass sie mit positivem Ergebnis möglich ist, da schon stärker verschiedene Formen aus der gleichen Verwandtschaftsgruppe (Helarctos x Melursus) Nachkommen miteinander haben können.)
Die Unterteilung der Braunbären in 3 Arten ist nur noch bei Mohr zu finden:
Ursus arctos
Ursus middendorfii
Ursus horribilis
Wo immer die anderen Autoren auf diese Formen zu sprechen kommen, schreiben sie:
Ursus arctos (arctos)
Ursus arctos middendorfii
Ursus arctos horribilis
Sollten alle Bärenformen der Familie Ursidae miteinander fertile Nachkommen hervorbringen können (die Untersuchungen sind noch nicht abgeschlossen), so sind auf genetischer Ebene alle Gattungen und Arten zu einer Art zusammenzufassen ( - was auch im Falle sekundärer Artbildung (s.o.) zutrifft); und wir würden schreiben:
Ursus arctos arctos (mit allen Formen)
Ursus arctos malayanus
Ursus arctos thibetanus
Ursus arctos ursinus
Ursus arctos americanus
Ursus arctos maritimus
Ursus arctos ornatus etc.
Bei Einbeziehung des Pandabären [vgl. O'Brien et al. 1985] sind genetisch vielleicht zwei Arten, nämlich
Ursus arctos und
Ursus melanoleuca
zu unterscheiden. Nach den vorliegenden zahlreichen Untersuchungen gehören jedenfalls so verschiedene Formen wie die Eis- und Braunbären genetisch sicher zur selben Art, was ebenso auf die Braun- und Schwarzbären zutrifft. Hier können wir auf festem Boden die Formen auf die Ebene des genetischen Artbegriffs bringen.
Auch bei den noch nicht gründlich durchforschten Artengruppen ist in der Regel kein so umfangreiches Arbeitsprogramm nötig, wie es Lamprecht mit den Bohnen durchgeführt hat (er musste ja auch mit einem möglichst umfangreichen Material arbeiten, um seinen neuen Ansatz wirklich abzusichern). Bei reziprok nicht verschiedenen F1-Bastarden entfällt in der Regel die Schwierigkeit eines so großen Aufwands, da alle bisher bekannten Beispiele, in denen bestimmte Merkmale reziprok nicht überführbar sind, in der F1 (je nachdem, welche Art als Mutter gewählt wurde) auch einen verschiedenen Phänotyp aufwiesen, d.h. Gleichformigkeit der F1 kann bei reziproken Kreuzungen als erstes Indiz für die Zugehörigkeit zur gleichen Art gewertet werden.
In diesem Zusammenhang sei an die Kreuzungen zwischen Pferd und Esel erinnert: Die Unterschiede zwischen Maultier (Mutter: Pferd) und Maulesel (Mutter: Eselin) sind beträchtlich. Es muss sich hier um (Oo-)plasmatische Unterschiede handeln, denn die Chromosomenstruktur ist in der F1 in beiden Fällen gleich. Interessanterweise berichtet Chandley (1981) von mehreren Fällen fruchtbarer Maultiere,*und die hier beschriebenen Erbgänge sind in weitgehender Übereinstimmung mit den Lamprechtschen Ergebnissen bei Pflanzen.
*Sie erwähnt Bielanski (1955), der alle zur Verfügung stehende Literatur zu diesem Thema seit 1527 ausgewertet hat und alles in allem "30 cases of alleged fertility in she-mules and 2 or 3 cases in she-hinnies" aufführt. "Fertile male hybrids have, however, never been reported".
"Old Bec" ist als eines dieser fruchtbaren, weiblichen Maultiere beschrieben worden und Chandley schreibt (1981, pp. 105/106):
Judging from a published photograph she appears to be a typical mule (L. Travis, personal communication), yet she produced two offspring. One was a typical she-mule sired by a donkey, who unlike her mother proved to be sterile, the other was a horse, described a being 'of good conformation', sired by a stallion. This son of 'old Bec' was not only fertile, producing a number of colts and fillies, but also served as a popular saddle horse around the area.
The types of progeny produced by 'Old Bec' are in fact consistent with all other reported cases from allegly fertile mules (Bielanski, 1972; Savory, 1970) in showing that when a female mule is sired by a donkey, the progeny is always mule or 'mule-like' and when sired by a horse, they are typical horse. Furthermore, the progeny of rare fertile female hinnies, when sired by a donkey are always donkey (Bielanski, 1955; Savory, 1970). (Matings of female hinnies to stallions appear not to have produced any outcome, but reports of such matings are extremely scarce). There is in fact such a consistency in the reporting of these matings that one is led to the same conclusion as Andersen (1939) that the ova of she-mules which produce viable pregnancies carry only horse chromosomes, while those of the hinny, carry only donkey chromosomes. In other words, viable pregnancies appear to arise from ova carrying only maternal chromosome sets, those carrying paternal chromosomes or the foetuses resulting from them, appear to be eliminated.
Als Mechanismus schlägt sie mit Michie (1953, 1955) und anderen Autoren Chromosomenaffinität vor und meint u.a. (p. 107):
'Affinity' could also explain the observed rapid reversion to parental type in interspecific hybrids (Wallace & Gunn, 1965) and may be of evolutionary significance in maintaining species integrity (Michie 1955).
One problem remains however...there is a total absence of the type of offspring expected if conception arose out of fertilization of a hybrid ovum containing its paternal chromosome set.
Die Frage nach der Bedeutung der (Oo-)plasmatischen Konstitution wird für das vorliegende Problem von der Verfasserin nicht gestellt. Auch die von ihr zitierten Autoren sind bei diesem Thema noch weitgehend von der Kernautonomie überzeugt und suchen die Erklärung in der Konstitution und im Verhalten der Chromosomen. Letztere spielen sicher bei dieser Frage eine Rolle - jedoch nicht unabhängig von der Konstitution des (Oo-)Plasmas. Die gewichtigen Unterschiede zwischen Maultier und Maulesel beweisen ja bereits die Bedeutung der plasmatischen Konstitution für die Genregulation. Wie schon betont sind die genetisch-chromosomalen Verhältnisse in der F1 in beiden Fällen gleich: diese Kernstrukturen können also den Unterschied nicht bedingen.
Könnten nicht cytoplasmatische Strukturen auch für die Regulation der Chromosomenverteilung bzw. der Chromosomenelimination verantwortlich sein? (Chandley weist darauf hin, dass Wodsedalek schon 1916 in seiner Arbeit über die Spermatozyten-Meiosis des Maultiers Fälle von Chromosomenelimination beschreibt, - was an die oben pp. 223, 232 zitierten Beispiele der Fisch- und Froschhybriden erinnert.)
Wäre es einfach eine Frage der Chromosomenaffinität, die aus einer "physical attraction of centromeres of similar ancestry" (Chandley, p. 107) resultierte (für welche Hypothese es meines Wissens bislang keine molekularen Daten gibt), dann müsste der männliche Chromosomensatz genauso häufig in der Eizelle auftreten wie der weibliche und damit eine entsprechende Nachkommenschaft möglich sein. Dafür gibt es nun allerdings nicht den geringsten Hinweis (vgl. das obige Zitat).
Aufgrund dieser Daten erscheint die von Michie 1955 diskutierte Alternativhypothese der 'Anziehung' durch ein Polelement der Zelle (die Spindel wird sowieso von den plasmatischen Strukturen gebildet) für den vorliegenden Fall wahrscheinlicher. Aber wir sprechen auch hier wieder von offenen Problemen, die erst noch gründlich erforscht werden müssen, bevor eine definitive Antwort möglich ist.
Lamprecht beschreibt (1966, pp. 385 - 387) die reziproken Unterschiede zur Kreuzung Pferd und Esel und gibt anschließend seine genetisch-plasmatische Interpretation des Phänomens (wegen der möglicherweise paradigmatischen Bedeutung des Beispiels möchte ich den ganzen Passus zitieren):
Seit altersher sind die reziproken Bastarde zwischen Pferd (Equus caballus L.) und Esel (Equus asinus L.) sehr gut bekannt. Es sind dies bei Verwendung des Esels als Mutter der Maulesel (Equus hinnus genannt) und mit dem Pferd als Mutter das Maultier (Equus mulus L.). Es handelt sich demnach um zwei Artbastarde.
Wenn dieselbe erblich einheitliche Pferderasse und eine entsprechend einheitliche Eselrasse zur Herstellung dieser beiden Bastarde verwendet werden (was gewöhnlich der Fall zu sein pflegt; z.B. in Poitou, Südrankreich), so ist zu sagen, daß beide Bastarde dieselbe genotypische Konstitution haben werden. Aber die beiden Bastarde unterscheiden sich stark und ganz typisch voneinander, auch ziemlich gleichgültig, welche Rassen der beiden Elternarten zu den Kreuzungen herangezogen werden.
Der Maulesel ist erheblich kleiner, er hat die längeren Ohren des Esels, aber vom Pferd den schlankeren und längeren Kopf, die starken Schenkel, den in seiner ganzen Länge behaarten Schwanz und überdies noch die wiehernde Stimme.
Das Maultier ist in seinem Körperbau nur wenig schwächer als das Pferd und erinnert auch in der Gestalt des Körpers stark an das Pferd. Die Kopfform ist abweichend, die Ohren sind länger als beim Pferd und der Schwanz ist nur an der Wurzel behaart. An den Esel erinnern auch die schmächtigeren Schenkel und die schmäleren Hufe. Das Maultier hat die Stimme des Eselvaters.
Werden nun Maultier und Maulesel, ohne auf ihre Herstammung Rücksicht zu nehmen, von dem Standpunkt aus betrachtet, den gegenwärtig viele, wenn nicht die meisten Genetiker hinsichtlich Artbegriff und Evolution einnehmen, so würde wohl zweifellos der Schluß gezogen werden, daß sich diese beiden "Arten" in einem ganzen Komplex, d.h. in einer größeren Anzahl von Genen unterscheiden müssen.
Die reziproke Kreuzung zwischen zwei erblich einheitlichen Rassen von Pferd und Esel beweist aber, wie schon erwähnt, daß sie dieselbe genotypische Konstitution besitzen müssen. Worauf beruht nun der große und stets vorhandene Unterschied und weshalb sind diese beiden Bastarde immer vollkommen steril *? - Nebenbei sei hier nur erwähnt, daß bei solchen Bastarden wohl eine gewisse Variation, vor allem hinsichtlich Färbung, vorkommen kann, was aber ausschließlich auf die zur Kreuzung verwendeten Elternrassen zurückzuführen ist. Die oben angeführten markanten Unterschiede werden hiervon jedoch in keiner Weise berührt.
Nun sind die Dominanzverhältnisse, wie erwähnt, in beiden Kreuzungsrichtungen verschieden. Ganz dasselbe konnte auch bei Artkreuzungen zwischen Pflanzen festgestellt werden, so z.B. bei Epilobium, Chrysanthemum und Phaseolus (s.o.). A priori könnte man annehmen, daß dieselben Gene sich in verschiedenem Plasma verschieden manifestieren. Die Ergebnisse meiner oben besprochenen Artkreuzungen und vor allem die Kreuzungen mit Exmutanten haben die Ursache dieser verschiedenen Manifestation intraspezifischer Gene wie folgt eindeutig klargelegt. Die Einführung eines Allels eines interspezifischen Gens in ein artfremdes Plasma kann die Manifestation einer ganzen Reihe von intraspezifischen Genen stark abändern. In besonders hohem Grade dokumentiert sich diese Wirkung in Kreuzungen mit der lathyroides-Exmutante von Pisum (s.o.). Das artfremde Allel i-lath bedingte im artfremden Plasma eine Änderung der Manifestation von wenigstens 15 intraspezifischen Genen. Und naturbedingte Spezies, d.h. Arten, die durch die Allele von interspezifischen Genen getrennt sind, dürften laut allen bisher vorliegenden Ergebnissen zu urteilen immer verschiedenes Plasma besitzen.
Mit Hinblick auf das oben und schon früher geschilderte Verhalten interspezifischer Gene erscheinen die Verhältnisse beim Maulesel und Maultier nun in ganz natürlicher und einfacher Weise erklärbar. Pferd und Esel unterscheiden sich nur in einem einzigen interspezifischen Gen für den somatischen Teil der Tiere. Für den generativen Teil kommt noch ein zweites auf dieses eingestelltes Allel hinzu (s.o.). Die Einführung des für das Soma des Pferdes arteigenen Allels des spezies-trennenden interspezifischen Gens bedingt im Eselplasma die Änderung der Manifestation einer Anzahl von intraspezifischen Genen, die zu den oben angeführten Merkmalen des Maulesels führt. Ganz Entsprechendes gilt für die Einführung des anderen Allels desselben interspezifischen Gens vom Esel in das Pferd. Es führt zu den für das Maultier charakteristischen Merkmalen.
Sowohl Maulesel wie Maultier scheinen immer vollkommen steril [siehe Anmerkung zu p. 260] zu sein. Die Ursache hierfür ist, daß bei der Reduktionsteilung die für die Synthese der Gene verantwortlichen Progene für das artfremde Allel nicht erneuert werden, wie dies z. B. die Phaseolus-Kreuzungen mit coccineus als Mutter ausnahmslos gezeigt haben. Bei Pflanzen wurde festgestellt, daß hiervon gewisse Ausnahmen vorkommen können, nämlich dann, wenn mit dem männlichen Kern etwas Plasma in die Zygote mitkommt, wie dies in Phaseolus-Kreuzungen mit vulgaris als Mutter hat beobachtet werden können. Das im Vergleich mit vulgaris viel größere Pollenkorn von coccineus scheint hierfür prädisponiert zu sein. Mit diesem Plasma kommen Stoffe mit, die für die Erneuerung der Progene unerläßlich sind. In umgekehrter Richtung mit coccineus als Mutter ist unter einem sehr großen Material auch nicht ein einziger solcher Fall aufgetreten (näheres s.o.).
Was für Maulesel und Maultier gilt, dürfte wahrscheinlich für das ganze Tierreich Gültigkeit besitzen. Leider liegen aber besonders auf diese Frage abzielende Untersuchungen kaum vor.
Alle bisher und schon recht zahlreichen Beobachtungen sprechen eindeutig dafür, daß die Reproduktion arteigener Allele von interspezifischen Genen, wie sie aus vom Plasma zur Verfügung gestellten Stoffen durch die Wirkung von Progenen synthetisiert werden, für die unüberbrückbare Barriere zwischen naturbedingten Spezies und höheren Kategorien verantwortlich sind. Und eine Entstehung neuer Arten kann daher auch nur durch einen diese Barriere überwindenden Schritt, eine weitere Differenzierung des Plasmas für die Produktion zweier neuer Progene und damit diesen entsprechenden interspezifischen Genen stattfinden.
*Lamprecht würde heute sicher gern die oben zitierten Ausführungen von Chandley und anderen zur vereinzelten Fortpflanzung von Maultieren zur Kenntnis nehmen, zumal sie in weitgehender Übereinstimmung mit seinen Phaseolus-, Chrysanthemum und Epilobium-Beispielen sind.
Zurück zur systematischen Frage: Es handelt sich im vorliegenden Fall auch auf der genetisch-Lamprechtschen Ebene um zwei verschiedene Arten. Die beiden Ausgangsarten sind zwar miteinander kreuzbar, aber gleichzeitig durch die Nichtüberführbarkeit der artentrennenden Merkmale durch die meiotisch-plasmatische Barriere absolut voneinander getrennt, d.h. durch die Kreuzungen wird niemals eine Rekombination der artentrennenden Merkmale stattfinden.
Ad 2. Wir müssen aber dennoch die Frage behandeln, ob es sich im vorliegenden Falle um sekundäre Arten handeln könnte. Dabei ist es sinnvoll, die anderen Equus-Arten in die Untersuchungen miteinzubeziehen, nämlich Equus przewalskii, E. burchelli (Burchells Zebra mit den verschiedenen Unterarten burchelli, antiquorum und boehmi), E. grevyi (Grévys Zebra), Equus zebra zebra und E. zebra hartmannae, E. quagga, E. hemionus (der asiatische Wildesel mit seinen verschiedenen Unterarten hemionus, hemippus, khur, kiang, onager) - vgl. dazu die Zusammenstellung von Gray 1971, pp. 94 - 113.
Bei den Equidae sind die Hybriden aus Artkreuzungen meist steril und aus den Unterartkreuzungen - wie zu erwarten - fertil. Zu den Ausnahmen von dieser Regel dürften die 'Artkreuzungen' zwischen den verschiedenen 'Zebraarten' gehören, aus Grays Zusammenstellung war jedoch nichts über die Fertilität der Zebrahybriden zu entnehmen. (In Klammern sei in diesem Zusammenhang noch ein ganz anderes Beispiel erwähnt: auch die verschiedenen Giraffenformen wurden zunächst als eigene Arten beschrieben. Nachdem sich jedoch herausstellte, dass alle untersuchten 'Arten' miteinander fertile Nachkommen hervorbringen konnten, wurde hier konsequent die tertiäre Nomenklatur durchgeführt.)
Die bei den Pferd-Esel-Kreuzungen gemachten Erfahrungen wiederholen sich in ähnlicher Weise bei anderen Artkreuzungen der Equidae. So ist Grays Arbeit z.B. zu entnehmen, dass die Hybriden aus der Kreuzung Equus asinus x E. burchelli in der Regel steril sind, "although fertile females may occasionally occur" (p. 94). Ähnliche Formulierungen finden wir außer für die schon diskutierten Pferd-Esel-Kreuzungen auch für die Hybriden von
E. asinus x E. burchelli antiquorum und
E. burchelli antiquorum x E. caballus.
Für weitere ähnliche Fälle gibt es Hinweise. Für die Hybriden aus
E. asinus x E. zebra und
E. burchelli boehnmi x E. caballus, d.h. für Hybriden mit
anderen Zebra-Unterarten als bei den zuvor erwähnten Kreuzungen, schreibt Gray (pp. 99 und 105): "Female hybrids have normal uteri, oestrous cycles and corpora lutei, but primordial germ cells are rare." Zur Kreuzung E. asinus taeniopus x E. hemionus hemippus lesen wir p. 101: "The hybrids are probably sterile." Und zur Kreuzung E. caballus x E. zebra (Gray führt in der Regel das männliche Tier zuerst auf) schreibt sie, dass die Meinungen über die Fertilität der weiblichen Hybriden auseinandergehen.
Durch unterschiedliche Chromosomenzahlen bei verschiedenen Arten wird zwar die Situation in einigen Fällen noch komplizierter, auf der anderen Seite hat E. caballus 64 Chromosomen und E. przewalskii 66 Chromosomen, dennoch sind die Hybriden in beiden Geschlechtern fertil (vgl. Gray, p. 107). Wir haben ja mit Herre und Röhrs schon auf ähnliche Fälle aufmerksam gemacht, vgl. auch p. 178. Für das Pferdebeispiel ist nach der genetischen Systematik E. caballus przewalskii zu schreiben.
Man muss sich bei den Kreuzungen der Equidae vor Augen halten, dass die Pferd-Esel-Kreuzungen aus wirtschaftlichen Gründen am häufigsten durchgeführt worden sind und dass es sich bei der Fertilitätsfrage um die am gründlichsten untersuchten Hybriden handelt. Obwohl also die anderen Daten auf zahlenmäßig geringeren Untersuchungen beruhen, zeichnet sich in dieser Tierfamilie dennoch ein genereller Trend ab, und man muss einfach fordern, dass dahinter eine allgemeine Gesetzmäßigkeit steht, die dem Lamprechtschen Ansatz nahekommt. Wenige Regulatorgene in Verbindung mit einer spezifischen plasmatischen Konstitution [Pferd und Esel scheinen sich u.a. in ihrem Mitochondriengenom zu unterscheiden] dürften für die Artunterschiede verantwortlich sein.
Interessant für die systematischen Fragen ist die Tatsache, dass die genetisch-Lamprechtsche Systematik mit dem status quo der Artunterscheidung im Falle der Equidae bereits in vielen Fällen kongruent ist, auf der anderen Seite jedoch noch eine Revision für das Przewalski-Pferd und die Zebras notwendig wird.
Zur oben gestellten Frage, ob es sich nicht auch bei den heutigen Arten der Equidae um sekundäre Arten handeln könnte, wollen wir zunächst hervorheben, dass wir in diesem Falle Arten annehmen müssten, die durch genetische Strukturausfälle alle ungefähr gleich weit voneinander entfernt sind. Das klingt derzeit wenig wahrscheinlich. Da aber zu dieser Frage bisher keine molekulargenetischen Daten vorliegen, wollen wir das Problem für eine endgültige Entscheidung zunächst offenlassen, aber für die systematische Frage vorerst von den derzeit wahrscheinlichsten Ergebnissen ausgehen.
Ähnliche Verhältnisse wie bei den Equidae scheinen sich z.B. bei den Felidae in der Gattung Panthera mit den Arten Panthera leo, P. onca, P. pardus und P. tigris wiederzufinden (reziproke Unterschiede und ab und zu Fertilität bei den weiblichen Hybriden).
Den Hauptpunkt zum genetisch-plasmatischen Artbegriff möchte ich hier noch einmal besonders hervorheben: Zahlreiche Untersuchungen haben gezeigt, dass das mütterliche Plasma (Eiplasma) auf die Expression des väterlichen Erbguts eingestellt sein muss, wenn ein lebensfähiger und fertiler Organismus aus der Verbindung zwischen mütterlicher und väterlicher Keimzelle entstehen soll. Immer wieder stellen wir fest, dass viele systematische Arten einer Gattung (sowohl in der Botanik als auch in der Zoologie) miteinander kreuzbar sind und fruchtbare Nachkommen haben können. Oft trifft das auch noch auf die Gattungen ganzer Familien zu (pp. 20, 23, 27). Zeigen jedoch verschiedene Arten grundsätzliche Unterschiede in ihrem Bauplan, dann kann aus einer solchen Verbindung nichts Vernünftiges mehr entstehen. Viele Taubenarten z.B. sind miteinander kreuzbar (wie pp. 23-27 im Detail aufgeführt). Würde man jedoch (durch künstliche Befruchtung) eine Taube mit einem Alkenvogel oder einem Papagei kreuzen, dann könnte daraus nichts Vernünftiges hervorgehen, obwohl es sich dabei nach Steinbacher, 1972/1982, um die "nächstverwandten" Vogelgruppen (eine Unterordnung und eine Ordnung) handelt. Warum geht das aber nicht? Warum könnten keine intermediären Zwischenformen entstehen und warum könnten die Merkmale nicht nach den Mendelschen Gesetzen in der 2. Filialgeneration frei kombiniert werden? (Man würde dann durch Rekombination ganz neue Gattungen und Familien erhalten!)
Der genetisch-plasmatische Artbegriff gibt darauf die eindeutige Antwort: Im Falle, dass die Taube die Mutter sein soll, ist das Ei- und Artplasma nicht in der Lage, die abweichenden, artfremden Gene des Papageienvaters "anzuschalten" und zu exprimieren. Soll der Papagei die Mutter sein, so ist das Ei- und Artplasma der Papageienmutter genausowenig in der Lage, die artfremden Gene des Täuberichs anzuschalten und zu exprimieren. Die befruchtete Eizelle (soweit eine Vereinigung der Keimzellen hier überhaupt noch möglich wäre - vielleicht auch nur mit zusätzlichem Eingriff des Menschen) - könnte wohl noch die kernunabhängigen Teilungen durchführen, die Ontogenese käme jedoch sehr bald ins hemizygote Stadium (nur noch das Erbgut der Mutter, also der haploide Chromosomensatz, würde exprimiert) und die Entwicklung fände schnell ein Ende.
Sehen wir uns einige unterschiedliche Merkmale zwischen beiden Vogelordnungen näher an:
|
Tauben |
Papageien |
|
1. relativ kleiner Kopf |
relativ großer Kopf |
|
2. schmaler Schnabel (ohne Gelenk am Oberschnabelansatz, Unterschnabel nicht schlittenartig beweglich) |
starker, gekrümmter Oberschnabel (Oberschnabelansatz mit Gelenk, Unterschnabel schlittenartig beweglich) |
|
3. Kropf produziert Kropfmilch (einen käseartigen Brei, der von der Schleimhaut des Kropfes geliefert wird) |
Kropf produziert keine Kropfmilch. Statt dessen wird im Kropf Nahrung mit Enzymen aus dem Drüsenmagen aufgeweicht. |
Vergleich zwischen Tauben- und Papageienschädel (aus Grzimeks Tierleben 1980):
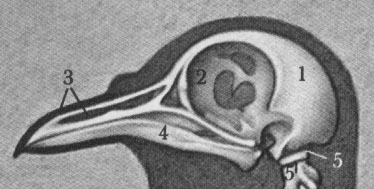
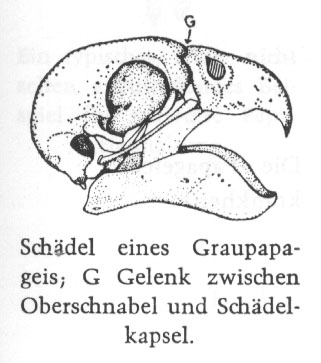
Taube (Ausschnitt): 1 Hirnschädel; 2 Augenhöhle (Orbita); 3 Gesichtsschädel, Grundlage des Oberschnabels; 4 Unterkiefer (Mandibula), Grundlage des Unterschnabels; 5 Erster Halswirbel (Atlas)
Zur Kropfmilch schreibt Johnston 1980, p. 237:
...unter dem Einfluß eines Hormons des Hypophysenvorderlappens, des Prolaktins (das gleiche Hormon beeinflußt bei Säugetierweibchen die Milchbildung!), verdickt sich die Schleimhaut während des Brütens gewaltig und stößt fettige Schichten in den Hohlraum der beiden Kropftaschen ab.
Dorst schreibt 1972, p. 105 (Das Leben der Vögel I):
Die Sekretbildung setzt bei beiden Geschlechtern am 8. Tag nach Brutbeginn ein und dauert bis zum 16. Tag nach dem Schlüpfen der Jungen. Mit der hochgewürgten Kropfmilch werden die Jungen geatzt. Dieser Fall ist bei den Vögeln einzigartig, die Tauben sind so die einzigen, die ihre Jungen wenigstens zum Teil mit einem selbsterzeugten Sekret ernähren.
Entsprechend verhalten sich die Jungvögel:
"Im Gegensatz zu vielen anderen Vogeljungen verlangt die junge Taube ihre Nahrung nicht durch Schnabelaufreißen, sondern wartet geduldig auf ihren Teil." (Dorst: Das Leben der Vögel II, 1972, p. 692)
Man könnte eine ganze Palette weiterer Merkmalspaare aufführen, in denen sich die Tauben von den Papageien (und umgekehrt) voneinander unterscheiden.
Dabei muss man sich vergegenwärtigen, dass bei jedem dieser Merkmalspaare das jeweils andersartige, artfremde Merkmal des Vaters im Ei- und Artplasma der Mutter nicht realisiert werden kann - auch nicht als intermediäre Erscheinung - weil hier die artfremden Gene und Gensysteme im Plasma der Mutter in der Regel gar nicht erst angeschaltet werden und damit auch nicht exprimiert werden können (vgl. Zitat Ohno, p. 224).
Hinzu kommt, dass eine intermediäre Ausbildung etwa des Oberschnabelgelenkes keinen Sinn gäbe (es handelte sich dann um ein nicht gebrauchsfähiges Gelenk). Beim Kropf könnte man sich allerdings eine Kombination von Kropfmilch und präparierter Nahrung vorstellen - es fragte sich dabei nur, ob sich die Enzyrne des Drüsenmagens mit der Milch vertragen würden. Wenn Kreuzung mit fertiler Nachkommenschaft zwischen solch verschiedenen Bauplänen möglich wäre, gäbe es in der F2 neben neuen Gattungen und Familien auch eine große Zahl in sich widersprüchlicher und nicht lebensfähiger Rekombinanten. Was sollte beispielsweise eine Taube mit beweglichem Oberschnabel ohne entsprechend ererbte Verhaltensprogramme, die diese Sondermerkmale beim Papagei erst nützlich machen? Wozu sollte ein Papagei "seine" Kropfmilch mit Enzymen aus dem Drüsenmagen aufweichen? Wenn auch neue Kombinationen denkmöglich sind, so macht doch die absolute Trennung der verschiedenen Vogelgruppen durch die genetisch-plasmatische Artbarriere allen Spekulationen ein Ende: Das mütterliche Plasma kann die artfremden Merkmale nicht exprimieren, hier weder intermediär in einem Hybridorganismus noch durch Rekombination in späteren Generationen. Interessanterweise gibt es auch paläontologisch keine Zwischenformen zwischen Tauben und Papageien, genausowenig wie zwischen Tauben und Alkenvögeln, so dass man die Tauben weder von Papageien noch von Alken (bzw. die Papageien von den Tauben und diese von den Alken) realgenetisch ableiten könnte. (Nach Wolters 1982, stehen die Taubenvögel mit den Pterocliformes jedoch zwischen den Pelecaniformes und Psittaciformes und die Aussage wäre entsprechend zu modifizieren.)

Abb. 44a: Hypothetisches Stammbaumdiagramrn der von Wolters angenommenen Ordnungen der Vögel.
Das Verhältnis der Länge der ausgezogenen Stridie zueinander deutet das Verhältnis der Länge der Zeiträume an, aus denen die einzelnen Ordnungen fossil nachgewiesen sind. Punktierte Linien bezeichnen zwei oder mehr mögliche Verbindungen.
(Aus: Wolters 1982)
Und wie steht es mit dem von Wolters (1982) u.a. postulierten gemeinsamen Vorfahren von Tauben und Papageien (vgl. Abb. 44a)? Wie sollte er beschaffen sein? Ihm mussten sowohl die charakteristischen Merkmale der Tauben als auch der Papageien fehlen. Und wie käme dieser vermutete ziemlich unspezialisierte Vorfahr zu den neuen Merkmalskomplexen dieser neuen Vogelordnungen? Wie soll sich z.B. das Oberschnabelgelenk in Tausenden von kleinen Mutationsschritten entwickelt haben? Ein Gelenk hat biologisch erst einen Sinn, wenn es funktioniert! Wie kamen die Prolaktinrezeptoren in den Kropf der Taube? Wie kam es zur genauen zeitlichen Abstimmung zwischen Kropfmilchproduktion und Brutverhalten? Wie kam es zur angemessenen Mengenproduktion der Kropfmilch? Alles nur Zufall? Auch die sinnvollen Verhaltensweisen müssen in den Problemkreis miteinbezogen werden. Denn was würde die Kropfmilch nützen, wenn die Tauben - statt ihre Jungen damit zu füttern - die Milch irgendwohin auswürgen würden, etwa wie die Eulen ihre Gewölle? Die uns bekannten richtungslosen Mutationen können die komplexen neuen Synorganisationen nicht erklären (vgl. pp. 325-438).
Zur Paläontologie schreibt Murton 1982, p. 936 über die Taubenvögel:
By the time they appear in the fossil record the Columbiformes are already so well differentiated that their phylogenetic relations cannot be determined with certainty.
Sowohl Tauben als auch Papageien sind aus dem Oligozän bekannt und hatten nach gängiger geologischer Zeitrechnung ihre charakteristischen Merkmale schon vor mehr als 26 Millionen Jahren voll ausgebildet! Das darf man wohl Konstanz der Merkmale nennen. Nur über die Zufallsevolution ist nichts bekannt.
Zur Frage nach den verwandtschaftlichen Beziehungen stellt Ziswiler in seinem Buch "Spezielle Zoologie - Wirbeltiere" 1976, p. 462, über die Papageien fest:
Es gelang bis jetzt nicht, überzeugende Beziehungen dieser Ordnung zu anderen Gruppen zu belegen.
- Was sich in den letzten Jahren genausowenig geändert hat wie in über hundert Jahren evotutionistischer Ableitungsversuche davor.
Man könnte eine Spezialarbeit allein über die Vogelordnungen und Familien anfertigen, die durchweg mit ähnlichen Ergebnissen das gleiche Prinzip demonstrieren würden (vgl. Abb. 44a). Und das gilt für das gesamte Tier- und Pflanzenreich!
Im Gegensatz zu den zitierten Ab- (und Um-)bauerscheinungen bei den sekundären Arten, sind die primären Arten durch den Aufbau [vgl. auch de Vries, zitiert p. 51] neuer anatomischer, physiologischer und ethologischer (Strukturen und) Systeme, die aufgrund der artplasmatischen Barriere bei Kreuzung prinzipiell nicht reziprok überführbar und homozygot zu exprimieren sind, absolut voneinander getrennt.
Ad 3. Die Frage nach der Zusammenfassung stark verschiedener und somit klar voneinander unterscheidbarer Organismen, die dennoch zur selben Art gehören, haben wir oben schon ausführlich diskutiert (vgl. pp. 196-204). Ich möchte an dieser Stelle nur noch einmal hervorheben, dass es ja gerade dieses Bewusstsein des realgenetischen Zusammenhangs von stark verschiedenen Formen ist, worauf der besondere Wert des genetischen Artbegriffs mit beruht. Wissenschaftliche Arbeitsweise verlangt den Abschied von Phänomenen als Artkriterien, wenn diese aus einer allgemeinen Gesetzmäßigkeit (hier der Neukombination der Gene) herzuleiten und gleichzeitig sowohl in der großen Mehrzahl aller Fälle in der Praxis als auch aus zahlreichen logisch-theoretischen Gründen nicht als artentrennend aufzufassen sind (vgl. pp. 79-112, 122-137, 170-204).
Durch den genetischen Artbegriff wird außerdem das ganze System des Pflanzen- und Tierreichs übersichtlicher und - last not least - sollte man bei der Frage nach dem Ursprung der Arten wissen, was man mit dem Artbegriff meint. Der Artbegriff sollte wissenschaftlich definiert sein. (Es ist einfach unwissenschaftlich in einem Fall Rekombinanten als Varietäten, im nächsten als Subspezies und im übernächsten als eigene Arten oder gar Gattungen zu beschreiben. Entweder sind alle Rekombinanten Arten oder keine. Als großer Vorteil erschien bisher jedoch die Möglichkeit, mit einem wissenschaftlich nicht definierten Artbegriff zahlreiche Beweise für 'die Entstehung der Arten' zu liefern, die sich nun zum größten Teil als Populationen Mendelscher Rekombinanten erweisen.)
Ad 4. Das kann nun tatsächlich ein Problem in der Kreuzungspraxis sein (vgl. pp. 170-194). In vielen Fällen ist jedoch eine Entscheidung über Kreuzungen mit weiteren nah verwandten Formen möglich (vgl. die obigen Kreuzungspolygone). Außerdem haben wir ausführlich dokumentiert, dass verschiedene Linien ein- und derselben Art steril, semisteril oder fertil bei Kreuzung mit Linien anderer Arten sein können (vgl. p. 127 dieser Arbeit), d.h. dass eine sekundäre Artbarriere durch Verwendung verschiedener Linien derselben Form nachgewiesen werden kann. Schließlich eröffnen sich durch die fortschreitende Molekulargenetik völlig neue Perspektiven zum Nachweis der Funktion von Nukleotidsequenzen durch Funktionsausfall (vgl. Weintraub et al. 1985 zum Thema ANTI-SENSE RNA AS A MOLECULAR TOOL FOR GENETIC ANALYSIS).
Die Frage 5 wurde mit den vorliegenden Bemerkungen schon weitgehend beantwortet. Zur Beschreibung der Variabilität der Formen an den verschiedenen Arealen leistet der morphologische Artbegriff nach wie vor gute Dienste und er ist ein wichtiges Übergangsstadium zu einem möglichst objektiv-naturwissenschaftlichen Artbegriff, dem wir mit der Lamprechtschen Definition ein gutes Stück näher gekommen sind. Den systematischen Status quo als endgültige Antwort auf die Frage nach einem naturwissenschaftlichen Artbegriff beizubehalten, ist aufgrund der pp. 62 - 66 zitierten Schwächen unmöglich. Systematiker sollten vielmehr in verstärkter Zusammenarbeit mit den klassischen und molekularen Genetikern die großen Aufgaben angehen, die noch einer Lösung harren.