F2-ZUSAMMENBRUCH, HYBRIDENSTERILITÄT UND HYBRIDENSTERBLICHKEIT UND -SCHWÄCHE (PUNKTE 4,2 UND 1, p. 68)
(1) F2-ZUSAMMENBRUCH
Wie oben p. 68 zitiert, sind die F1-Hybriden normal, kräftig und fertil, aber die F2 enthält viele schwache oder sterile Individuen.
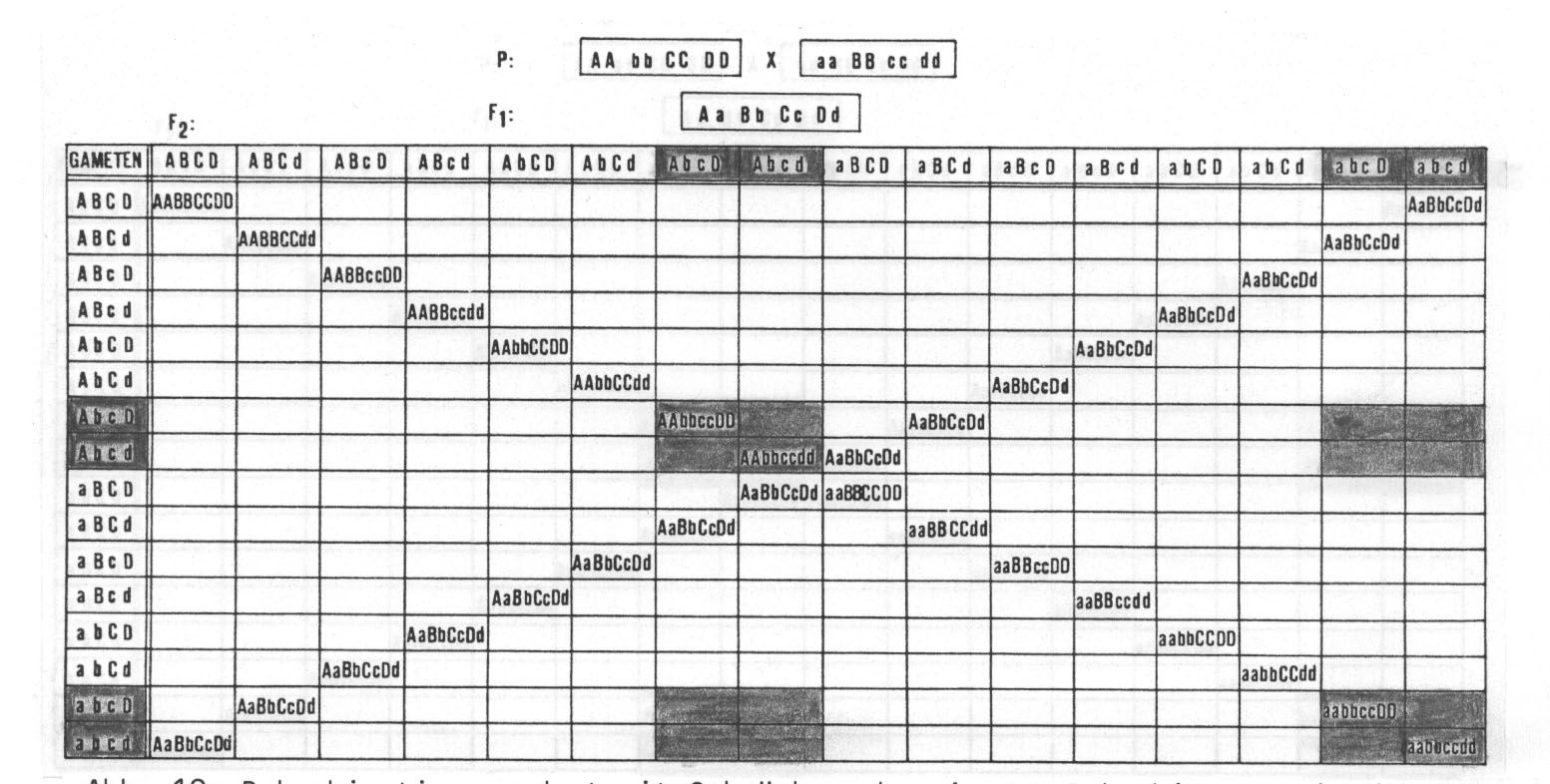
Reicht dieser Punkt aus, um bei den Elternlinien genetisch von zwei verschiedenen Ausgangsarten sprechen zu können? Sehen wir uns unser Rekombinationsquadrat zur F2-Spaltung noch einmal unter einem anderen Gesichtspunkt genauer an. Wir sind bei der bisherigen Betrachtung davon ausgegangen, dass die rezessiven Gene a, b, c und d weder als einzelne noch in Kombination eine deutliche Schwächung oder gar Sterilität der Individuen zur Folge hatten - wofür es Hunderte von Beispielen gibt. Auf der anderen Seite können rezessive Gene als einzelne oder in Kombination im homozygoten Zustand aber auch Schwächung bis zur Sterilität bedeuten.
Abb. 19: Rekombinationsquadrat mit Schwächung bestimmter Rekombinanten durch epistatische Geninteraktionen.
Nehmen wir für unser Rekombinationsquadrat an, dass die Genkombination bb cc (unabhängig von den Genen A und D und der Frage, ob die Letzteren im dominanten oder rezessiven Zustand vorkommen) Schwächung der betroffenen Individuen bedeutet, dann ist davon schon ein Sechzehntel aller oben im Rekombinationsquadrat gegebenen Rekombinanten betroffen (in der Abbildung mit grauen Flächen gekennzeichnet).
Zur Frage, was rezessive Gene molekularbiologisch sind, schrieb Watson 1976, p. 190 (siehe auch Watson et al. 1987, p. 222):
Most mutant genes are recessive with respect to wild-type genes. This fact, puzzling to early geneticists, is now partially understood in terms of the gene-enzyme relation. The recessive phenotype often results from the failure of mutant genes to produce any functional protein (enzyme). (Kursiv vom Verfasser)
Kaudewitz erklärt die Rezessivität am Beispiel der rot- und weißblühenden Erbsen-Pflanzen 1983, p. 187 (r steht für weiß):
Das Allel r kennzeichnet eine Mangelmutante. Es vermag nicht mehr die Synthese des roten Blütenfarbstoffs der Erbse zu induzieren. Der betreffende Genort kann verloren gegangen oder derart verändert sein, daß keine Transkription seiner genetischen Information mehr erfolgt. Auch eine Unterbindung der Translation ist möglich. In beiden Fällen entsteht kein enzymatisch wirkendes Polypeptid mehr. Hat die Mutation jedoch nur zu Veränderungen eines oder weniger Aminosäurereste geführt, dann wird zwar noch anstelle des Wildenzyms ein homologes Polypeptid translatiert, ihm fehlt, wie das völlige Ausbleiben der Färbung der Blüten der weißblühenden Rasse zeigt, jegliche Enzymaktivität.
(Weitere
Ausführungen bei Kacser und Burns 1981, Fincham 1983, Karlson
1984).
Auf Details möchte ich später zu sprechen kommen.
Bei Haustieren, Kulturpflanzen und genetischen Versuchsorganismen wie Drosophila, der Bierhefe etc. ist es ohne Frage passend, von 'wild-typ genes' und 'Wildenzymen' in Abgrenzung zu den mutierten bzw. veränderten Genen und Enzymen zu sprechen. Da es jedoch auch bei Wildtyp-Genen Dominanz und Rezessivität gibt, kann man für diesen Fall auch ganz allgemein von Genen und Enzymen sprechen. Weißblütige Wildformen beispielsweise gibt es in zahlreichen Pflanzenfamilien. Obwohl oben im Prinzip richtig beschrieben, sei an dieser Stelle noch darauf hingewiesen, dass die Anthozyanbildung vom Zusammenspiel zahlreicher Gene abhängt. Bei der Erbse z.B. sind 6 - 8 Gene für die Ausbildung der normalen roten Blütenfarbe nachgewiesen worden (vgl. Lamprecht 1974; Einzelheiten zur Anthozyansynthese bei Heß 1981, 1983). (Vgl. auch p. 193)
Beim Zusammenkommen mehrerer rezessiver Gene muss man sich vor Augen halten, dass 'das Zusammenspiel der genetisch gesteuerten Leistungen eines Lebewesens einen sehr hohen Grad des Zusammenwirkens seiner zahlreichen Einzelkomponenten aufweist' (Kaudewitz p. 198). Daraus folgt, dass beim Zusammenkommen mehrerer funktionsloser Enzyme ein additiver Effekt auftreten kann, der mit Schwächung und/oder Sterilität der Nachkommen von Hybriden verbunden ist. Im Prinzip reichen dafür schon zwei rezessive Gene im homozygoten Zustand (in anderen Fällen sogar im heterozygoten Zustand; vgl. p. 124) aus, nämlich in dem Augenblick, in dem sie in einer Rekombinante eine wichtige Schlüsselfunktion bei den komplexen Biosynthesen eines Organismus beeinträchtigen. Dobzhansky hat übrigens schon 1955 und 1970 ein Modell epistatischer Interaktion für Nachteile im heterozygoten Zustand vorgeschlagen (Einzelheiten bei Kimura 1983).
Je mehr unterschiedliche Einzelkomponenten in verschiedenen Linien ausfallen (und hier gerade noch kompensiert werden können), d.h., je mehr funktionslose Polypeptide gebildet werden, desto größer wird die Wahrscheinlichkeit, dass die F2 zahlreiche schwache oder sterile Individuen enthält. Gehen wir nur einmal davon aus, dass in unserem Rekombinationsquadrat alle Genotypen, die homozygot in den rezessiven Genen a b, a d, b c und c d sind, auch geschwächt sind (dreifach homozygote sollen davon noch weiter in Mitleidenschaft gezogen, aber noch schwach fertil sein, vierfach homozygote seien steril). Dann sind davon bereits etwa ein Fünftel betroffen (von 256 sind es 47, also 209 normale und 47 schwächliche Individuen, was einem Verhältnis von 4,45 : 1 entspricht. Von den 47 schwächlichen Rekombinanten sind 34 doppelt homozygot, 12 dreifach homozygot und nach unserer Definition stark geschwächt und eine vierfach homozygot und damit steril; vgl. Abb. 20).
Kreuzt man nun - soweit das bei den schwach fertilen möglich ist - die dreifach homozygot-rezessiven Individuen miteinander, z.B. AA bb cc dd x aa BB cc dd, so erhält man in der F2 bereits ein Sechzehntel sterile Rekombinanten (6,25 %) und 37,5 % sind schwach fertil; dafür sehen dann die restlichen 56,25 % besser aus.
Sind jedoch die beiden dreifach homozygot-rezessiven Eltern in dem verbleibenden Genort heterozygot (Aa bb cc dd X aa Bb cc dd), dann findet man bereits in der F1 ein Viertel sterile Individuen (die Hälfte ist dann dreifach homozygot-rezessiv und ein Viertel zweifach homozygot-rezessiv; die dominanten Gene liegen nur im heterozygoten Zustand vor).
Man kann sich das für die verschiedensten Möglichkeiten in allen Einzelheiten weiter ausrechnen (z.B. auch für den Fall, dass bestimmte Genkombinationen von 2 Genen Sterilität zur Folge haben) - die entscheidende Frage, die sich hier anschließt, lautet: Handelt es sich beim Zusammentreffen solcher Linien und Formen um zwei verschiedene Arten?
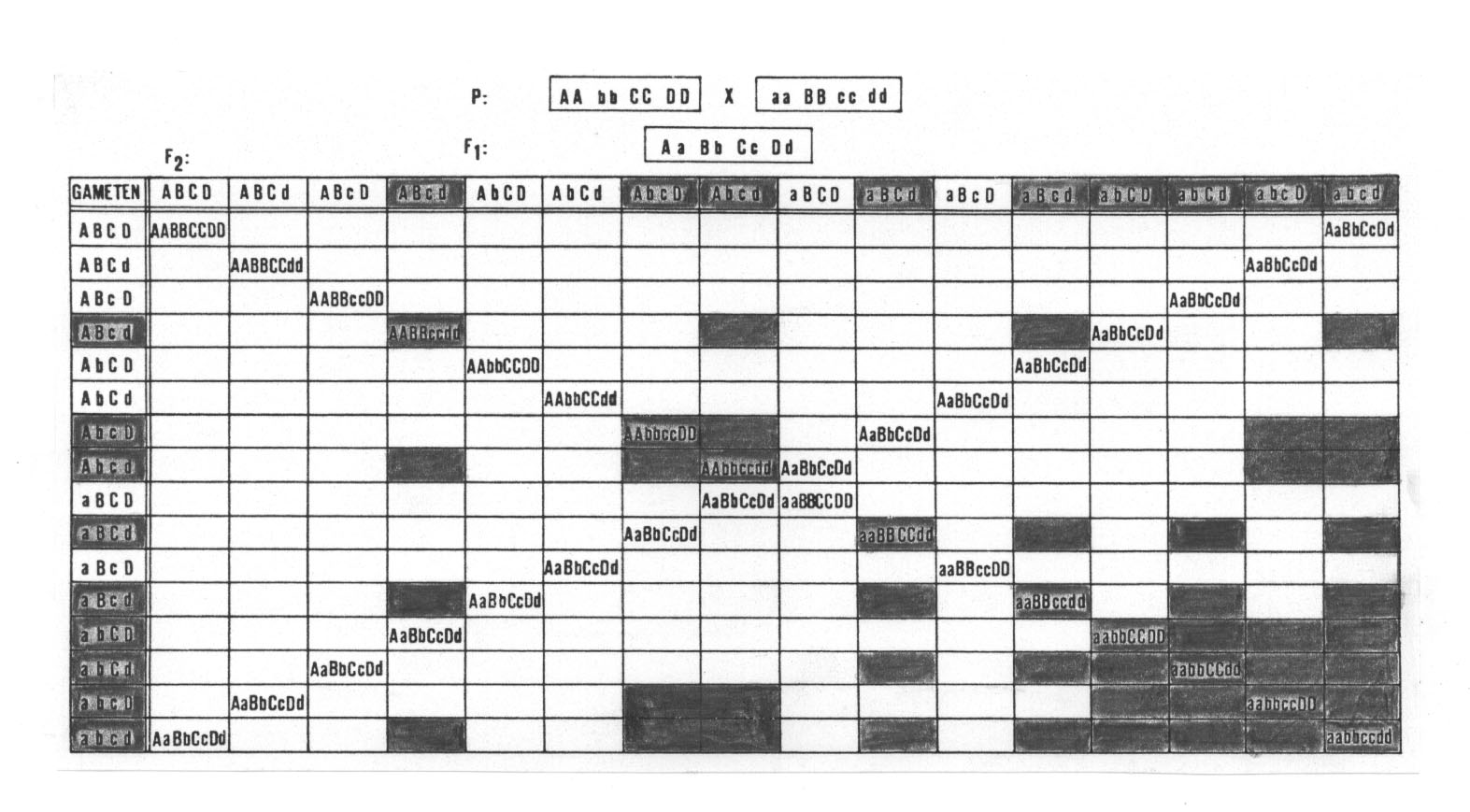
Abb. 20: In dem
vorliegenden Rekombinationsquadrat sollen alle Genotypen, die homozygot in
den rezessiven Genen a b, a d, b c, und c d sind, auch geschwächt
sein (das soll jedoch nicht auf a c und b d zutreffen). Dreifach homozygote sollen noch weiter in Mitleidenschaft gezogen, aber noch schwach fertil sein, vierfach homozygote seien steril. Fazit: 18,36 % sind geschwächt (alle grauen Felder).
Kreuzt man die dreifach homozygot-rezessiven Eltern miteinander, so
erhält man in der F2 bereits ein Sechzehntel sterile
Rekombinanten und 37,5 % sind schwach fertil.
Sind jedoch die dreifach
homozygot-rezessiven Eltern in dem verbleibenden Genort heterozygot (Aa bb
cc dd x aa Bb cc dd), dann findet man bereits in der F1
ein Viertel sterile Individuen.
Weitere Details im Text.
Man könnte argumentieren, dass wir jetzt tatsächlich einen genetischen Isolationsmechanismus vorliegen haben, der noch mit verschiedenen präzygotischen Isolationsmechanismen kombiniert sein kann.
Gelingt es einem jedoch, von der homozygoten Rekombinanten AA BB CC DD (die in der F2 und den folgenden Generationen immer wieder herausspalten wird) eine Population zu erhalten, dann ist diese Population mit beiden Ausgangspopulationen voll fertil! Die Isolation ist dann durch Rekombination aufgehoben worden. Die Reversibilität würde gegen das Aufstellen zweier Arten sprechen.
Die Tatsache, dass bei Kreuzung zweier "Arten" häufig nur ganz bestimmte Linien aus diesen Arten miteinander kreuzbar sind, andere dagegen nicht, unterstreicht die Problematik: Mit der Grenzziehung der genetischen Isolation als Arttrennungskriterium würde es sich bei den miteinander fertilen Linien zweier "Arten" im Grunde genommen doch nur um die Repräsentanten derselben Art handeln, bei den anderen, nicht miteinander fertilen Linien aber um Angehörige verschiedener Arten - was machen wir nun mit den Linien ein und derselben Ausgangsart, die solche unterschiedlichen Fertilitätsgrade mit anderen "Arten" aufweisen? Hier würde ja der Artbegriff mitten durch "die Art" gehen! (Lamprecht z.B. beschreibt solche Fälle für Phaseolus und Chrysanthemum 1966.)
Bevor wir weiter auf diese Fragen eingehen, wollen wir einige Beispiele im Sinne der oben aufgeführten Rekombinationsquadrate geben.
Grant berichtet 1963, pp. 388/389 (ähnlich 1976):
The F1 hybrid between two species of tarweeds, Layia gaillardioides and L. hieracioides (Compositae), is vigorous and semifertile, but 80 percent of the F2 individuals are weak and many of these are dwarfish or lethal. Similarly, many F2 progenies derived from interspecific crosses in Gilia include a fair to large proportion of weak, dwarfed, or lethal types. In the F2 of the semifertile hybrid Gilia leptantha X G. latiflora, for example, about half of the individuals are as vigorous as the parents and the other half are weak or dwarfed. In Layia, Gilia, and other plants an interspecific F2 generation often exhibits continuous variation from fully vigorous individuals through plants slightly smaller or weaker than normal to inviable types.
Dobzhansky schreibt 1977, pp. 178/179 über die männlichen und weiblichen Hybriden der Kreuzung Drosophila pseudoobscura X D. persimilis:
Depending on the strains crossed, the full number of chromosomal bivalents (five) are formed in the primary spermatocytes, or some of the chromosomes remain unpaired. Regardless of complete or incomplete pairing, the cell-division machinery grossly malfunctions, and nothing like normal spermatozoa are formed. However, female hybrids between the same Drosophila species are fertile and can be backcrossed to both parental species.
...Genic sterility results from failures of the gene complements of distinct species to function harmoniously in the hybrids. Although no disharmonies may arise in the somatic cells, the processes of germ-cell formation may be disrupted. In such a case the hybrid is somatically vigorous but sterile.
Genic sterility is known in hybrids between species with very similar or even identical gene arrangements.
Nach Hinweis, dass genische Sterilität häufiger bei Tieren und chromosomale Sterilität häufiger bei Pflanzen auftritt und dass die interspezifischen Hybriden unterschiedliche Fertilitätsgrade aufweisen können, wird noch einmal der anfangs erwähnte Punkt hervorgehoben:
The degree of hybrid breakdown varies depending upon the strains of the parental species used to produce hybrids.
Zu den Beispielen Grants wäre noch nachzutragen, dass bei Gilia genische und chromosomale Sterilität in Kombination auftreten können.
Handelt es sich nun in solchen Fällen um eindeutig verschiedene Elternarten? Bei den Hybriden von Drosophila pseudoobscura X D. persimilis sind nur die Männchen steril, die Weibchen aber mit beiden Elternarten fertil. Wendet man die Artdefinition aufgrund genetischer Isolation konsequent auf einen solchen Fall an (weitere Beispiele unten), dann beweisen die hybriden Männchen, dass es sich um zwei verschiedene Arten handelt, die hybriden Weibchen jedoch, dass man es mit ein- und derselben Art zu tun hat - was nun auch nicht gerade Klarheit in die Fragestellung bringen würde.
Weitere Beispiele: Bei der Kreuzung Chironomus thummi thummi X Ch. th. piger sind bei bestimmten Linien (im Gegensatz zu dem oben zitierten Drosophila-Fall) nur die weiblichen Hybriden steril. Hägele berichtet (1984, p. 111):
Sterility in both sexes of the Ch. th. thummi H1 (female) X Ch. th. piger E (male) crosses is stock-specific. Crosses in the same direction, but with different thummi and piger stocks show sterility only in the female hybrids (Hägele, in prep.). Sterility is in all the cases the consequence of rudimentary or non-developed gonads.
Der Autor weist u.a. darauf hin, dass bei der Kreuzungsrichtung Ch. th. piger (weibl.) X Ch. th. thummi (männl.) die Hybriden durch Chromosomenaberrationen und verschiedene Entwicklungsstörungen gekennzeichnet sind, wogegen in der Regel bei reziproken Bastarden beide Geschlechter steril sind.
Durch die tertiäre Nomenklatur hält der Autor für die verschiedenen Chironomus-Linien den Status der Unterart aufrecht, doch ist er mit Miehlbradt und Neumann (1976) der Meinung, dass es sich hier um eine "intermediate position between species and subspecies" handelt (p. 105). Nach neodarwinistischen Arttrennungskriterien müssten die miteinander nicht voll fertilen Linien Artstatus erhalten.
Das Phänomen einer Hybridzone, in der der Genaustausch stark variiert, so dass der Genfluss in einigen Gegenden gering ist, in anderen aber weite Populationen beherrscht, wird von Harrison (1983) beschrieben (pp. 245/246):
The field crickets Gryllus pennsylvanicus and G. firmus hybridize and/or overlap in a zone that stretches from the BIue Ridge Mountains in Virginia to southern Connecticut (Fig. 1). Mixed or intermediate populations (determined by electromorph frequencies at an Esterase locus and by distributions of morphological characters) have been found (1) along the Blue Ridge Mountains and in the Shenandoah Valley in Virginia, (2) at the northern end of the Blue Ridge in Maryland, (3) in western New Jersey, (4) at the southern end of the Hudson Valley in New York, and (5) in southern Connecticut (Harrison and Arnold, 1982; See Fig. 1 for localities). The extent of hybridization appears to vary geographically. In Virginia and Maryland, mixed populations of the two eggoverwintering, fall adult Gryllus species consist mostly of crickets that can be classified as one of the two parental types. Presumably, little gene exchange has occurred. In contrast, examination of electrophoretic and morphological characters in mixed/intermediate populations from New York and Connecticut suggests substantial hybridization and introgression.
Seine Studien zur Fertilität der Gryllus-"Arten" fasst der Autor folgendermaßen zusammen (pp. 250/251):
Crosses between G. pennsylvanicus males and G. firmus females consistently failed to produce offspring. In contrast, the reciprocal cross resulted in viable and fertile F1 hybrids. The outcome was not dependent on the locality from which the parents were obtained. Observations of spermatophore transfer demonstrated that the failure of the G. pennsy1vanius male X G. firmus female cross was indeed the result of a post-mating barrier.
Die Kreuzung Gryllus pennsylvanicus (weiblich) X G. firmus (männlich) brachte lebensfähige und fertile Nachkommen hervor, die reziproke Kreuzung nicht. An einigen Stellen scheint der Genfluss gering zu sein, an anderen Stellen umfasst er große Populationen. - Um nicht je nach Kreuzungsrichtung und Breite der Hybridzone zu widersprüchlichen Schlussfolgerungen zu kommen, sollte der Artbegriff weiter gefasst werden.
Wie ist nun der folgende Fall nach neodarwinistischen Arttrennungskriterien zu beurteilen? Trpis, Perrone, Reissig und Parker berichten über die Kreuzbarkeit verschiedener Arten des Aedes scutellaris Komplexes 1981, p. 313 (abstract):
Most of the interspecific crosses in Aedes scutellaris complex result in undirectional cytoplasmic incompatibility. The case of cytoplasmic incompatibility presented here involves Aedes polynesiensis and Ae. kesseli. While the cross Ae. kesseli (female) and Ae. polynesiensis (male) is compatible, the reciprocal cross is not. By treating the larval stages of the parental stock with heat or the antibiotics tetracycline and chloramphenicol, the incompatible cross Ae. polynesiensis (female) and Ae. kesseli (male) is made compatible. Both sexes of the hybrid progeny were fertile and no hybrid breakdown occurred in any subsequent generation tested up to F9. ...
Die Barriere wurde wahrscheinlich durch bestimmte Mikroorganismen vom Rickettsien-Typus verursacht. (Weitere Beispiele aus der Literatur: Siehe die Arbeit der zitierten Autoren sowie J. A. Breeuwer und J. H. Werren 1990. Microorganisms associated with chromosomal destruction and reproductive isolation between two insect species. Nature 346, 558 - 560 (1990))
Bei konsequenter Anwendung der obigen Arttrennungskriterien, handelt es sich beim Vorhandensein solcher (cytoplasmatische Sterilität verursachender) Mikroorganismen um zwei verschiedene Arten, bei Abwesenheit um Linien derselben Art. Darüber hinaus kann man je nach Behandlung die eine Art in die andere überführen bzw. aus einer Art zwei machen.
Die Frage wird noch delikater, wenn man sich vergegenwärtigt, dass die interspezifischen Bastarde auch beim Aedes scutellaris Komplex unterschiedliche Fertilität aufweisen können.
Dev und Rai schreiben (1984, p. 83):
A comparison was made of karyotypes of 5 species in the Aedes scutellaris group and their hybrids.
(p. 89:) The chromosomal rearrangements ... did not by themselves fully explain the differences in the fertility of interspecific hybrids reported earlier (Dev & Rai, 1982). Often the type of hybrid and the backcross parent determine their fertility.
Aedes malayensis, Ae. alcasidi, Ae. polynesiensis, Ae. pseudoscutellaris und Ae. kesseli wurden von den Autoren untersucht.
Hilburn und Rai dokumentieren einen weiteren Fall für die Kreuzung von Ae. aegypti und Ae. mascarensis 1982, p. 59:
Perhaps the closest relative of Ae. aegypti which is the species Ae. mascarensis, found on the island of Mauritius. Successful hybridization of these species in the laboratory was reported by McClelland and Mamet with fertile progeny produced by both reciprocal matings. However, the subsequent work of Hartberg and Craig revealed that some hybrid crosses produced many abnormal sons, indicating hybrid breakdown. Of particular interest was the observation that abnormal development of the sexual phenotype occurred in male progeny produced by backcrosses involving Ae. aegypti females and F1 males obtained by crossing Ae. aegypti females with Ae. mascarensis males.
Die Anwendung des neodarwinistischen Artbegriffs würde auch bei diesen Beispielen zu den oben zitierten Schwierigkeiten führen: Je nach Aedes-Linie, Geschlecht des Bastards, Art des Rückkreuzungs-Elters sowie An- und Abwesenheit bestimmter Mikroorganismen käme man zu verschiedenen Ergebnissen.
Wir stoßen im übrigen hier auch wieder auf das Phänomen der Umkehrbarkeit oder Aufhebbarkeit der Schranke zwischen den "Arten". Ich hatte bei der Diskussion der präzygotischen Isolationsmechanismen schon wiederholt darauf aufmerksam gemacht, dass wegen verworrener Konsequenzen in zahlreichen Fällen, alle umkehrbaren Isolationsmechanismen als Arttrennungskriterien entfallen sollten. Das trifft in gleicher Weise auf die postzygotischen Isolationsmechanismen zu. Kawanishi und Watanabe berichten über die Kreuzung Drosophila melanogaster X D. simulans 1981, p. 1128:
Unfortunately, Wood and Ringo (1980) were only able to produce interspecific hybrid males carrying the X chromosome of D. simulans, since the X chromosome of D. melanogaster is lethal to hybrid males. Recently, Watanabe (1979) discovered a mutant of D. simulans, Lethal hybrid rescue (Lhr), which renders viable hybrid males between D. melanogaster females and D. simulans males, even though these hybrid males have the X chromosome of D. melanogaster.
Die Umkehrbarkeit spricht gegen ein solches Arttrennungskriterium. Andernfalls könnten zwei Arten durch eine oder wenige Mutation(en) zu einer Art verschmelzen. (Zur Fertititat bedarf es eines weiteren Schritts. Vgl. P. Hutter und M. Ashburner (1987): Genetit rescue of viable hybrids between Drosophila melanogaster and its sibling species. Nature 327, 331 - 333. (Interessanterweise sind jedoch Hybridweibchen aus den Kreuzungen D. simulans, D. mauritiana und D. seychellia bereits fertil!))
Werfen wir einen Blick auf das Pflanzenreich: Bei Anwendung solcher postzygotischer Isolationsmechanismen könnte man auf die Idee kommen, dass selbststerile Fremdbefruchter nur artfremden Pollen produzieren, - was wohl kein Biologe behaupten wird. (Überdies kennen wir Fälle, in denen die Selbststerilität durch Punkt- und Genommutationen aufgehoben worden ist - Campbell und Lacey 1982, Qaracéen 1982, Straub 1939, weitere Hinweise bei Gottschalk 1976.)
Um all die oben aufgeführten Schwierigkeiten und Widersprüche zu vermeiden, sollten solche Linien und Formen trotz der schwachen und/oder sterilen Individuen in der F2 (oder auch F1) als Angehörige ein und derselben Art betrachtet werden.
Da, wie schon oft hervorgehoben, die Synthetische Evolutionstheorie mit ihrem Artbegriff auf die Entstehung aller Arten und Lebensformen hinaus will, ist der oben behandelte genetische (postzygotische) Isolationsmechanismus für dieses Ziel denkbar ungeeignet. Denn es handelt sich bei dieser Barriere in der Regel um die Degeneration von Strukturen - Gene und Enzyme haben ihre Funktion verloren, was bei Addition in Rekombinanten zu Schwäche und/oder Sterilität führen kann. Der Ursprung völlig neuer funktionsfähiger Strukturen und Systeme, die zum Aufbau der Komplexität der Organismen notwendig sind, kann schlecht mit dem Ausfall wichtiger Strukturen erklärt werden.
Wir wollen der Frage nach dem Ausfall von Genen und Enzymen noch weiter nachgehen, denn es wäre ja denkbar, dass die in unseren Rekombinationsquadraten (pp. 122, 125) zunächst noch notwendigen dominanten Gene unter bestimmten Voraussetzungen ebenfalls ihre Funktion verlieren können:
Positionseffekte, Suppressor-Mutationen, durch weitere Rekombination veränderter genetischer Background, Gen-ökologische Veränderungen etc. können den Ausfall bestimmter Genwirkungen kompensieren (die Organismen verfügen über zahlreiche Möglichkeiten, ihr physiologisches Gleichgewicht wieder zu normalisieren). Die F1 - falls überhaupt noch eine gebildet wird - ist dann völlig steril (Beispiele bei den Kreuzungspolygonen pp. 170-193), und der postzygotische Isolationsmechanismus ist perfekt.
Spätestens an diesem Punkt würden wohl die meisten Biologen mit Klotz (vgl. Zitat p. 121) die Frage, ob es sich jetzt endlich um neue Arten handelt, bejahen.
Es sprechen jedoch mehrere biologische Tatsachen dagegen, die mir so gravierend erscheinen, dass sie für unsere Fragestellung unbedingt noch miteinbezogen werden müssen.