DER ARTBEGRIFF BEIM RASSENKREIS DER KOHLMEISE UND BEI DER SILBER- UND HERINGSMÖWE
In den meisten neueren Arbeiten zur Evolutionstheorie sowie in bald allen zoologischen Lehrbüchern werden entweder die Kohlmeise oder die Silber- und Heringsmöwe oder beide Rassenkreise zusammen als Musterbeispiele für Artbildung durch geographische und ethologische Isolation aufgeführt. Stresemann und Timophéeff-Ressovsky haben schon in den Titeln ihrer Originalarbeiten zum Larus-argentatus-Komplex von "Artbildung in geographischen Formenkreisen" (1947, 1959) gesprochen. Vogel und Angermann fassen die Bedeutung der geographischen in Verbindung mit der ethologischen Isolation für die Artbildungs-Frage unter Aufführung beider Rassenkreise wie folgt zusammen (1984, p. 505):
Sympatrische Arten oder Rassen können nur schwerlich aus einer einzigen Population entstehen, solange noch Panmixie herrscht. Andererseits kommt es bei allopatrischen Populationen, zwischen denen durch räuml. Trennung die Fortpflanzungsgemeinschaft aufgehoben ist, sehr leicht zu Artbildung, weil sie nun ihren autonomen spezif. Wirkungssystemen der Evolutionsfaktoren unterliegen. Daher konnte dieser Isolationsmechanismus bei fast allen Tiergruppen und versch. Pflanzengruppen nachgewiesen werden: Die vorhandene Artenfülle muß überwiegend auf ihn zurückgeführt werden.
Geograph. Rassen und Arten unterscheiden sich meist in mehreren Genen voneinander. Damit sind sie insgesamt für Anpassungen in verschiedener Richtung geeignet und besitzen daher große evolutionäre Potenzen. Die Unterscheidung "Rasse" oder "Art'' ist oft dadurch erschwert, daß die Verbreitungsgebiete der einzelnen Rassen einer polytypischen Art sich mosaikartig aneinanderfügen, und sich an den Arealgrenzen durch Bastardierung Übergangsformen entwickelt haben. Gerade solche Rassenkreise werden aber in den Fällen zu Beweisen der Speziation aus geograph. isolierten Rassen, wo sich extreme Glieder bzw. schon lange Zeit isolierte Rassen des Rassenkreises in einem sek. Überschneidungsgebiet unvermischt wie zwei neue Arten verhalten.
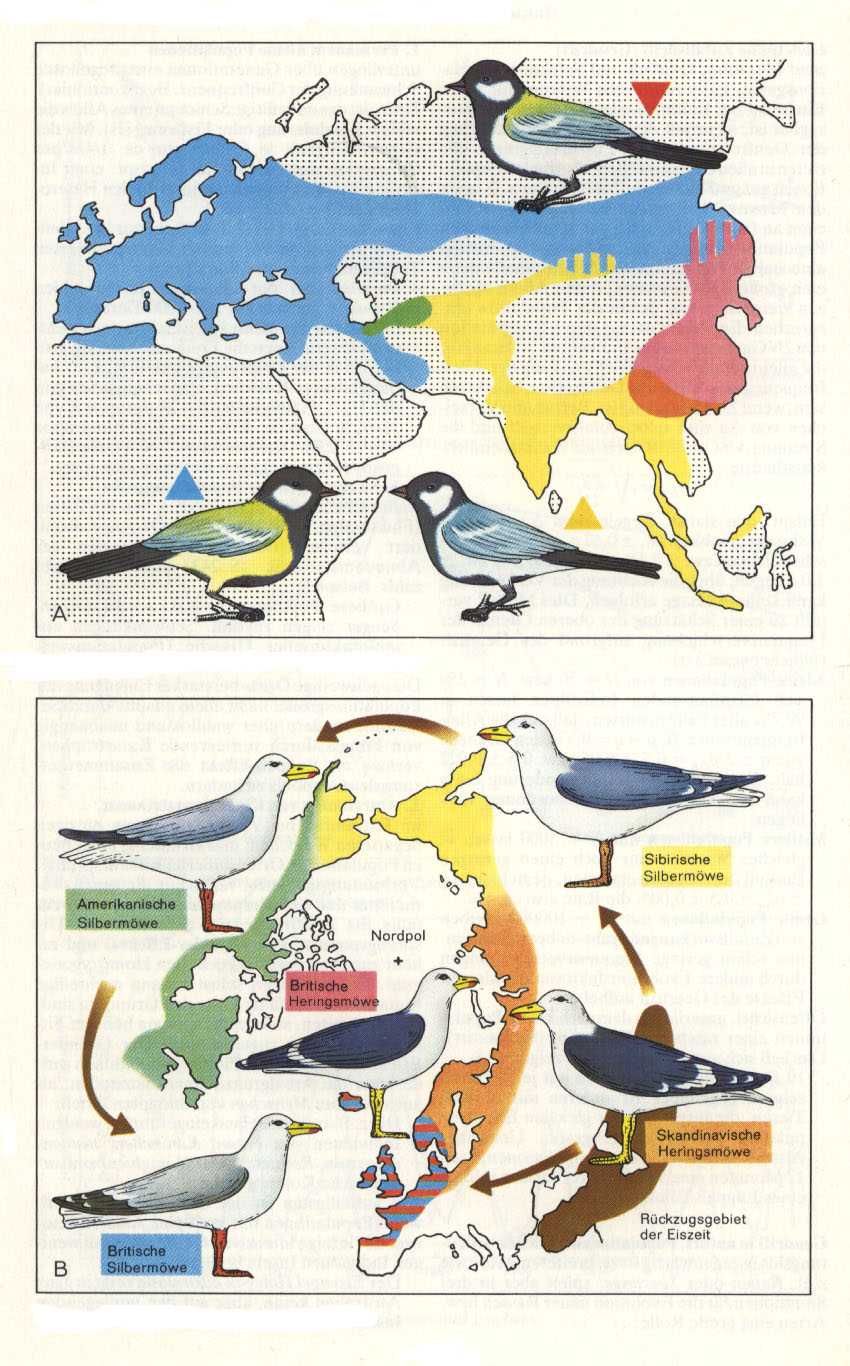
- Die Kohlmeise (Parus major) tritt in Eurasien in drei allopatrischen Formen auf. Die südliche Form geht in Persien kontinuierlich in die westliche, in Südostasien in die östliche infolge durchgehender Vermischung über. Nach der Eiszeit konnte die Westform so weit nach Osten vordringen, daß sie sekundär mit der Südform in Zentralasien und mit der Ostform im Amurgebiet sympatrisch wurde, ohne daß Vermischung auftritt (A).
- Nach der Eiszeit entwickelten sich aus Möwen der kaspischen Region die atlant. Britische Heringsmöwe (Larus fuscus graelsii), die Skandinavische Heringsmöwe (L. fuscus fuscus) und die pazifische Sibirische Silbermöwe (L. argentatus vegae), die sich in Nordamerika zur typ. Amerikanischen Silbermöwe (L. argentatus smithsoniatus) weiter entwickelte und schließl. als Britische Silbermöwe (L. argentatus argentatus) Nordwesteuropa besetzte. Sie lebt trotz des gemeinsamen Genpools rund um den Nordpol unvermischt mit der Britischen Heringsmöwe sympatrisch. (B).
In diesen Fällen war die Entw. eines Isolationsmechanismus möglich durch die Verzögerung des Genflusses durch eine lange Serie von Populationen (Distanz-Isolation)
Gut isolierte Gebiete (Inseln, Höhlen) beherbergen oft mehrere eng verwandte Arten, deren Stammart im Ursprungsraum einheitl. geblieben ist: Die Erstbesiedler aus der Stammpopulation waren in der Isolation bereits soweit genet. abgewandelt, daß eine Vermischung mit späteren Folgebesiedlern der gleichen Stammart unterblieb. (Kursiv von den Verfassern.)
Wir wollen uns an dieser Stelle nicht mit dem Thema Eiszeit als Verbreitungsursache auseinandersetzen (zum Thema Eiszeit gibt es die unterschiedlichsten Auffassungen, vgl. z.B. Daly 1972; Krause 1978, 1986; Oard 1984, 1985), sondern unser Augenmerk auf die geographischen Gegebenheiten
Abb. 11:
A: Rassenkreis der Kohlmeise (Parus major)
B: Circumpolare Überlappung bei Möwen.
Beide Abbildungen aus Vogel und Angermann 1984, Originale in Farbe.
Diskussion der Angaben im Text. - Die Silbermöwe, Larus argentatus, nistet übrigens auch auf der zum Naturschutzgebiet erklärten Ile de Port Cros (Mittelmeer, Südfrankreich), und in Übereinstimmung mit den Verbreitungsangaben von Tuck und Heinzel (1978) gibt Bologna (1980, p. 221) auch Nordafrika an.
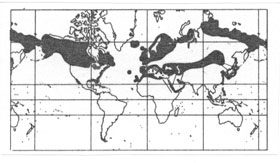
Abb. 12 : Verbreitung der Silbermöwe nach Tuck und Heinzel 1978. (Beachte besonders auch die mittelasiatische Verbreitung dieser Möwenart.)
und die ethologische Isolation richten. Sehen wir uns zunächst den Larus-argentatus-Komplex näher an. Ursprünglich wurde behauptet, dass es sich hier um ein ganz kontinuierliches, von allen Übergangsformen bewohntes, zirkumpolares Verbreitungsgebiet handelt, und dieser Gedanke taucht auch in neuerer Literatur noch auf (z.B. Jacobs 1969; Kull 1979; Remane, Storch und Welsch 1981 u.a.). "Bei nahezu allen früher beschriebenen kreisförmigen Überschneidungen scheinen die Dinge aber doch nicht so schematisch zu liegen wie ursprünglich geglaubt wurde. Dies gilt z.B. für den klassischen Fall kreisförmigen Überlagerns, bei dem zwei sympatrische Arten der großen europäischen Möwen, Larus argentatus und L. fuscus als Endglieder einer Kette von Unterarten längs der nördlich gemäßigten Zone siedeln. Hier haben Stresemann und Timophéeff (1947) gezeigt, daß vollständige Kontinuität der Populationen nicht vorhanden ist. Es sind häufige Lücken in der Reihe der Bevölkerungen zwischen Westeuropa und Ostasien da, und einige von ihnen sind soweit ausgeprägt, daß die Autoren sich veranlaßt sahen, den Larus-argentatus-Komplex in drei Arten aufzugliedern" (Mayr 1967, p. 401).
Hier reichte also die geographische Isolation unterscheidbarer Formen allein schon aus, mehrere Arten aufzustellen.
Der entscheidende Punkt für die Aufgliederung in mehrere Arten ist jedoch für die meisten Autoren die ethologische Isolation. Wo immer sich zwei Möwenpopulationen treffen und in der Regel nicht miteinander hybridisieren, spricht man von verschiedenen Arten. Wie in dem obigen Zitat von Vogel und Angermann (l984) heißt es in den meisten Besprechungen dieses Beispiels, dass sich aufgrund der mangelnden Paarungsbereitschaft die verschiedenen Formen wie neue oder echte Arten verhalten. Über die beiden Formen Laurs argentatus und L. fuscus schrieb Jacobs nach Hinweis, dass in der Kontaktzone keine Vermischung mehr auftritt (1969, p. 126): "Diese Endpopulationen stellen zwei echte Arten dar, sie sind reproduktiv voneinander isoliert, sie haben getrennte Gen-Poole."
Zwei weitere Punkte wollen wir in diesem Zusammenhang hervorheben: Mehrere Autoren erwähnen, dass die hier vorliegende ethologische Isolation keine absolute ist. Die Populationen dieser beiden Möwenformen verbastardieren sich "nur ganz selten" (Mayr 1967). "... gelegentlich können einzelne Bastardindividuen gefunden werden" (Lüers, Sperling, Wolf 1974) u.a.
Als zweiten Punkt möchten wir unterstreichen, dass sich die Hybriden durch ungestörte Entwicklung und volle Fertilität auszeichnen; - es gibt also keine echte genetische Barriere zwischen beiden Formen. Aus dieser Sicht ist der Hinweis zahlreicher Autoren, dass 'in diesen Fällen die Entwicklung eines Isolationsmechanismus durch die Verzögerung des Genflusses durch eine lange Serie von Populationen möglich war' (vgl. oben Vogel und Angermann für Meise und Möwe) wenig verständlich. Ohne Rücksicht auf die Frage nach der Kreuzbarkeit der verschiedenen Linien und der Fertilität der Bastarde, taucht dieser Gedanke bei den verschiedensten Autoren immer wieder auf. "Die entstandene biologische Isolation ist noch nicht absolut, sie hat das Stadium der genetischen Isolation im engeren Sinne noch nicht voll erreicht, denn gelegentlich können einzelne Bastardindividuen gefunden werden; damit wird unterstrichen, daß diese neuen Arten den status nascendi kaum erst überschritten haben, und zudem läßt sich auch hier ganz allgemein der Weg der Artbildung über die geographische Rassenbildung erkennen (Lüers, Sperling, Wolf). (Siehe auch Nachtrag p. 533)
Die entstandene biologische Isolation hat mit der eigentlichen genetisch-postzygotischen Isolation nichts zu tun. Die gegebene, unvollständige Isolation ist eine reine Verhaltensfrage: Wie oben zitiert, können selbst so minutiöse Unterschiede, wie die Färbung der Iris des Auges und die einer unbefiederten Hautpartie um das Auge "optische Artkennzeichen" abgeben, wie das für einige arktische Möwenarten nachgewiesen ist (vgl. Osche). Nach den jetzigen Kenntnisstand spielen Lernverhalten oder Prägung die entscheidende Rolle bei dieser Isolation. Wenn eine weitgehende Unterbrechung des Genflusses allein schon neue Arten ergäbe, dann müsste man auch beim Bernhardiner und Pekinesen von verschiedenen biologischen Arten sprechen. Dasselbe träfe auf das oben zitierte Beispiel der weißgefiederten und bläulich dunkelgrauen Form der Schneegans zu, wobei das letztere Beispiel durch die ethologische Barriere dem Möwenbeispiel entspricht.
Zur Farbe der Iris und des Augenrings als "Artkennzeichen" bei arktischen Möwen erwähnte Osche des Weiteren, dass bei Änderung der Farbe des Augenrings durch Bemalung sich bereits gebildete Paare wieder auflösen, da der geschminkte Partner offenbar nicht mehr als Artgenosse erkannt wird.
Ohne Berücksichtigung der genetischen Verhältnisse könnte ein konsequenter Vertreter der ethologischen Isolation als Arttrennungsfaktor darauf hinweisen, dass sich die beiden Partner jetzt wie zwei echte Arten verhalten. Das Verhalten allein reicht jedoch weder durch eine künstliche Modifikation noch durch eine Mutation oder Rekombination in der Iris- und Augenfarbe zur Trennung in zwei verschiedene Arten aus. Werden Möwenjunge auf 'artfremde' Eltern 'geprägt', dann ist die Grenze schon überwunden - abgesehen davon, dass in der Natur sowieso Bastardierung vorkommt. Statt "Artkennzeichen" sollte man besser von Populationskennzeichen sprechen. Die solcher Art gekennzeichneten Populationen selbst sollte man alle zusammen zu einer einzigen Art zusammenfassen und zur Unterscheidung der Populationen konsequent die tertiäre Nomenklatur gebrauchen.
Tegelström, Jansson und Ryttman haben bei Larus argentatus, L. fuscus und L. marinus etwa 40 verschiedene Loci auf Isozymdifferenzen untersucht und berichten 1980, p. 117:
There is identity or parallel variation for all loci investigated within the three species, making it impossible to distinguish between individuals from different species. This is in agreement with other genetic methods previously used to distinguish Larus species.
(p. 121:) The isozyme differences between species are often a useful tool in taxonomic determination. However, in this case, the three Larus species are not only identical for around 30 loci, they also seem to have the same alleles in the varying loci. This means that it is impossible to distinguish individuals of the three species by means of the 40 loci we have investigated. There is a striking similarity not only between the two species L. argenatus and L. fuscus but between these and the more distantly related L. marinus.
This is in agreement with other methods we have used to measure the genetic distance between L. fuscus and L. argenatus (Ryttman et al. 1978, 1979 and 1980). There seems to be little genetic differentiation of chromosomes, serum proteins, tissue structural proteins and enzymes in these species.
Und dann folgt ein Schluss, der allen bis dahin aufgeführten Tatsachen widerspricht:
There is, however, no question about the taxonomic status of the investigated species - they are definitely different species.
Warum gehören diese Formen, obwohl sie sich weder in ihren Isozymmustern noch in ihren Chromosomen, Serumproteinen, Gewebestrukturproteinen und Enzymen unterscheiden und darüber hinaus miteinander fertile Nachkommen haben können, weiterhin 'eindeutig verschiedenen Spezies' an? Die Antwort lautet wieder: weil man sie voneinander unterscheiden kann (Und selbst hier kann es noch zu Schwierigkeiten kommen. So berichten Tuck und Heinzel (übersetzt, bearbeitet und ergänzt von E. und F. Goethe) 1978 p. 136 über die Silbermöwe (die normalerweise einen grauen Mantel und rosig-fleischfarbene Füße hat): "In ihrer geographischen Verbreitung zeigen gewisse adulte Silbermöwen einen etwas dunkleren Mantel und gelbe Füße.'' Schwarzer Mantel und gelbe Füße aber gehören zu den Schlüsselmerkmalen der Heringsmöwe! Alles deutet darauf hin, dass hier Merkmale mendeln.) und sie sich in der Regel so verhalten.
Tatsächlich aber handelt es sich weder um 'neue Arten, die den status nascendi kaum erst überschritten haben', noch überhaupt um neue Arten. Drei Punkte können wir in diesem Zusammenhang festhalten:
1. Die geographische Verbreitung der verschiedenen Möwenrassen ist diskontinuierlich.
2. Die Isolation ist nicht genetisch, sondern rein ethologisch bedingt.
3. Die ethologische Isolation ist unvollkommen und reversibel.
Selbst einige evolutionistisch stark engagierte Forscher sind bei diesem und ähnlichen Beispielen skeptisch geblieben. Willmann weist (1985, p. 48) bei der Diskussion des Möwen-Beispiels auf Remane (1927), Hull (1965) und Klausnitzer und Richter (1979) hin, die (mit den Worten der letzteren Autoren) auch dann, wenn "Individuen zweier Populationen (A, B) nicht direkt miteinander fertile Nachkommen zu erzeugen vermögen ... sie trotzdem ... zu einer Art gezählt werden müssen, wenn beide Populationen mit einer dritten (C) fertile Nachkommen erzeugen können".
Willmann ist zwar der Auffassung, dass bei solchen Formenkreisen eine "Artaufspaltung" schon weit fortgeschritten sei (was jedoch oben mit zahlreichen Fakten in Frage gestellt wurde), kommt dann aber ebenfalls zu dem Schluss (pp. 48/49):
Solange aber die Aufspaltung nicht abgeschlossen ist, solange existiert auch keine absolute Fortpflanzungsisolation. Und solange dies nicht der Fall ist, besteht die Gesamtheit der Populationen eines solchen Formenkreises als einheitliche Biospezies.
Zum nächsten Beispiel, der geographischen Verbreitung und Artbildung bei der Kohlmeise, schreiben Remane, Storch und Welsch (1974, p. 261/262).
... die Rassenkreise der Kohlmeise, die sich in Ost- und Mittelasien überlappen, werden heute oft nur noch als Beispiel für unvollständige Artbildung angesehen. Sie zeigen aber das Prinzip der sich überlappenden Rassenkreise deutlich.
Sterben in einem solchen überlappenden Ring die mittleren Subspezies aus, so sind aus einer Art zwei geworden.
Zum letzten Satz ist festzustellen, dass im Falle eines Aussterbens der mittleren Subspezies, die beiden anderen genauso fruchtbar miteinander wären wie zuvor und dass sich dadurch selbstverständlich auch nichts an der normalen Entwicklung und Fertilität der Hybriden ändern würde. Es würde sich genetisch nach wie vor um eine einzige Art handeln. Andernfalls machen wir reversible geographische Verhältnisse und Zufälligkeiten sowie Lernverhalten zum Maßstab unseres Artbegriffs: Paaren sich zwei Populationen nicht, dann handelt es sich um zwei echte Arten, paaren sie sich aber an anderer Stelle doch, handelt es sich nur um eine Art. Gibt es genügend Zwischenformen bei einer weitverbreiteten Population, handelt es sich um eine Art, entfällt aufgrund irgendwelcher Umstände (sagen wir durch Veränderung der ökologischen Situation in einem bestimmten Gebiet) eine mittlere Subspezies, dann handelt es sich um zwei Arten. Gelingt es jedoch, die mittlere Subspezies als Rekombinante aus den anderen Subspezies wieder zu erhalten und in ihrem ursprünglichen Gebiet neu einzubürgern, verschmelzen die beiden 'Arten' wieder zu einer Art. Bleiben durch weitere Umstände nur die mittlere und die beiden 'äußersten' Subspezies erhalten, dann handelt es sich um drei Arten usw. Kann sich bei etwa gleicher geographischer Verbreitung die eine Art auch bei veränderten ökologischen Bedingungen mit all ihren Subspezies halten, bleibt sie die einzige Art, hat aber eine andere Art mehrere größere Populationsverluste unter gleichen Umständen, dann zerfällt sie in mehrere neue Arten, obwohl die genetische Divergenz zwischen den Endpopulationen der beiden Arten etwa gleich sein kann. In letzter Konsequenz heißt das, dass Verarmung des Genpools - Ausfall von Rekombinanten - Vermehrung der Artenzahl bedeutet.
Um solche und ähnliche Schlussfolgerungen zu vermeiden, sollten sämtliche reversiblen Isolationsmechanismen für den Artbegriff entfallen!
Wenn wir aber zeigen möchten, wie neue Arten im Sinne des herkömmlich-systematischen Artbegriffs entstehen, d.h. unter welchen Voraussetzungen Systematiker neue Arten unterscheiden, dann bieten solche Beispiele sicher eine überzeugende Erklärung. Die Synthetische Evolutionstheorie wollte mit ihrem populationsgenetischen Ansatz aber gerade den morphologischen Artbegriff überwinden. Statt dessen erklärt sie mit ihren Methoden oft nur, wie die Arten des herkömmlichen Artbegriffs entstehen und impliziert dabei durch Extrapolation in Zeit und Raum, dass das im Prinzip genau die Methode ist, wie die gesamte Organismenwelt ins Dasein gekommen ist.
Durch Verabsolutierung der einzelnen Isolationsmechanismen übertrifft der Neodarwinismus im Aufstellen neuer Arten sogar noch den morphologischen Artbegriff: Jedes der oben genannten Kriterien (verschiedene Biotopbesiedlung, jahreszeitliche und ethologische Isolation) wird für sich allein schon für ausreichend erachtet, um neue Arten zu bestimmen. Und das geschieht selbst da noch, wo kein morphologisch arbeitender Systematiker mehr verschiedene Arten unterscheiden könnte. Ich möchte beim Problem der Zwillingsarten auf diesen Punkt zurückkommen.
Kommen wir zum Rassenkreis der Kohlmeise zurück. Kühn und Heß schreiben 1984, p. 271:
Die geographischen Rassen einer Art bilden miteinander einen Rassenkreis oder eine Rassenkette. Vielfach sehen wir, daß benachbarte geographische Rassen einander ähnlich und die entfernteren Glieder der Rassenkette sehr stark voneinander verschieden sind. Ja, der Unterschied der äußersten Glieder kann sogar so weit gehen, daß diese keine Fortpflanzungsgemeinschaft mehr bilden, wenn sie zusammentreffen. Als Beispiel diene der Rassenkreis der Kohlmeise (Parus major). Er zerfällt in drei große Gruppen, geographische Hauptrassen, die jeweils wieder kleinere Teilrassen umfassen. Die europäischen Rassen haben grünen Rücken und gelbe Bauchseite (major-Gruppe); die Rassen von Persien an ostwärts bis zu den Sundainseln (bokharensis-Gruppe) besitzen keinen gelben Farbstoff; sie sind graurückig und weißbäuchig. Die Rassengruppe von Japan und China (minor-Gruppe) hat einen gelbgrünlichen Fleck am oberen Ende des sonst grauen Rückens. Überdies unterscheidet sich diese Ostrasse auch durch ihre geringere Größe von den übrigen. Die drei Hauptrassen treffen in drei Gebieten (Iran, Südchina und Amurgebiet) zusammen und bilden dort Mischbevölkerungen (Abb. l3).
In den zentralen Gebieten findet sich jeweils nur eine geographische Rasse; in den Randgebieten lassen sich die Individuen nicht eindeutig einer der Großrassen zuordnen.
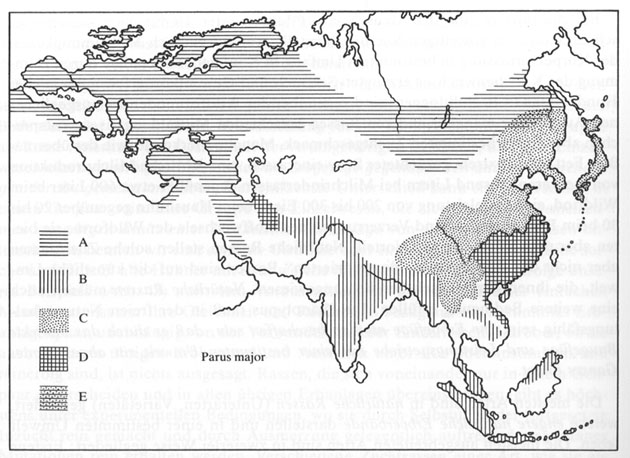
Abb.13: Verbreitung der Rassenkreise der Kohlmeise
Parus major. A, B, C Verbreitung der Hauptrassen, D, E
Vermischgebiete.
Aus Kühn und Heß 1984 nach Delacour, Vaurie und Mayr.
Die Zergliederung einer Art in geographische Rassen kann bei vollständiger räumlicher Trennung und hinreichender Dauer der Isolierung in die Trennung von Arten übergehen. Wenn die verwandten Arten später wieder in einem Gebiet zusammentreffen, verhalten sie sich wie artfremde zueinander. In Gefangenschaft können sie sich noch paaren, wie Nachtigall und Sprosser, die in einem gemeinsamen Wohngebiet zwischen Weichsel und Oder unvermischt nebeneinander leben.
Ich habe die einleitenden Worte von Kühn und Heß zu diesem Beispiel mitzitiert, weil es da heißt, dass der Unterschied der äußersten Glieder eines Rassenkreises 'sogar soweit gehen kann, daß diese keine Fortpflanzungsgemeinschaft mehr bilden, wenn sie zusammentreffen. Als Beispiel diene der Rassenkreis der Kohlmeise (Parus major)'.
Aber gerade so verhalten sich die Rassen der Kohlmeise nicht. 'Die drei Hauptrassen treffen in drei Gebieten zusammen und bilden dort Mischbevölkerungen.'
Im übrigen haben Remane, Storch und Welsch ihre oben zitierte Behauptung, dass hier das Prinzip der sich überlappenden Rassenkreise deutlich gezeigt wird, in der vierten Auflage ihres Lehrbuchs (1981) fallen gelassen. Mayr schrieb schon 1963/1967 p. 402: "In der Amurregion wurden sie früher als Beispiel zirkulärer Überlappung angesehen, aber sie bilden nach neueren Forschungen eine Bastardpopulation."
Aufgrund dieser Tatsachen erscheint mir das Beispiel der Kohlmeise noch schwächer als der Rassenkreis von Silber- und Heringsmöwe. Zum letzteren und zum Beispiel von Nachtigall und Sprosser möchten wir die Frage stellen, welchen Stellenwert eine Artbarriere hat, die jeder Sachverständige mit etwas züchterischem Geschick allemal wieder aufheben kann.
Ich kann Mayr nicht folgen, wenn er im Anschluss an das Möwen- und Meisenbeispiel behauptet (pp. 402/403):
Es ist unwesentlich, ob diese Beispiele für zirkuläres Überlagern einfach und schematisch sind oder sich etwas komplizierter darstellen, wie die neuen Untersuchungen an Larus argentatus zeigen. In jedem Falle kann der Prozess der geographischen Speziation Schritt für Schritt verfolgt werden. Einen schlagenderen Beweis für geographische Speziation gibt es wohl nicht.
- oder wenn er einleitend zu den kreisförmigen Überschneidungen auf p. 400 feststellt:
Eine vollkommene Demonstration der Speziation stellt eine Kette ineinander übergehender Unterarten dar, die eine Schleife oder einen übergreifenden Kreis bilden, dessen Endglieder sympatrisch geworden sind und sich nicht mehr kreuzen, obwohl sie durch eine vollständige Reihe ineinander übergehender oder hybridisierender Populationen miteinander verbunden sind. So sind diese Fälle beiläufig auch eine vortreffliche Illustration der "Artbildung durch Distanz". Die Entwicklung isolierender Mechanismen vollzog sich hier durch Hemmung des Genflusses über eine sehr lange Kette von Populationen. Obgleich nur einer der vielen Beweise für geographische Speziation, hat kreisförmiges Überschneiden immer als besonders überzeugend gegolten.
Wesentlich überzeugender erscheint mir ein Kommentar Goldschmidts zu diesem Thema aus seinem klassischen Werk THE MATERIAL BASIS OF EVOLUTION (1940, p. 168):
Some taxonomists claim that the existence of artenkreise demonstrates the gradation of the subsubspecies via subspecies into species at the end-points of the subspecific range, and even of those into subgenera and genera. I think that such conclusions amount to self-deception produced by mistaking nomenclature for evidence of macroevolution.
_________________________
NACHTRAG ZU SEITE 91 (BASTARDIERUNG ZWISCHEN LARUS ARGENTATUS UND LARUS FUSCUS)
Dass die beiden Rassen selten miteinander bastardieren liegt daran, dass sie unterschiedliche Brutplätze besetzen. Siedeln jedoch fuscus-Individuen in argentatus-Kolonien, dann liegt der Prozentsatz nach Zählungen aus den letzten 60 Jahren auf drei Nordseeinseln bei immerhin 6 Prozent! (Das Verhältnis beträgt 234 L. fuscus-Paare zu 30 Mischpaaren auf einem Hintergrund von etwa 106 100 argentatus-Paaren) (persönliche Mitteilung unter eigenen Beobachtungen und Quellenangaben von Herrn Dr. F. Goethe, Vogelwarte Helgoland, 1986).
Bleibt noch nachzutragen, dass auch Stegmann in seiner Larus-Monographie von 1934 die heute wieder zu einzelnen Arten erhobenen Populationen bereits zu einer Art zusammengeführt hatte. Der Grund, solchen Populationen dennoch Artstatus zu geben, wird von Stresemann und Timopheéff-Ressovsky u.a. wie folgt beschrieben (1947, p. 68): "...(es wird dadurch die beginnende und schon ganz reelle Formen annehmende Artdifferenzierung innerhalb dieses Formenkreises besonders unterstrichen."
Stegmann hatte jedoch die auf 5 Arten verteilten 19 Unterarten des Formenkreises zu einer Art zusammengefasst.
B.K. Stegmann (1934): Über die Formen der großen Möwen und ihre gegenseitigen Beziehungen. J. Ornithol. 82
E. Stresemann und N.W. Timophéeff-Ressovsky (1947): Artentstehung in geographischen Formenkreisen. 1. Der Formenkreis Larus argentatus-cachinnans-fuscus. Biol. Zentr. 66, 57 - 76
______________
NACHTRAG 2001: Bernd Petri ist bei seinen gründlichen Studien im Amurgebiet zur Frage DIE KOHLMEISEN (Parus major) - KONTAKTZONE AM MITTLEREN AMUR - EXISTIEREN AKUSTISCHE ISOLATIONSMECHANISMEN? (80 pp.; Diplomarbeit 1998, Fachbereich Biologie der Johannes Gutenberg-Universität Mainz) ebenfalls zum Ergebnis gekommen, dass "keine wirklich effizienten ethologischen und insbesondere keine akustischen Isolationsfaktoren" existieren. "Die Kohlmeisen im Gebiet beherrschen die, im Grunde sehr unterschiedlichen Dialekte von minor- und major-Kohlmeisen. Ebenso sind keine ökologischen Isolationsfaktoren zu erkennen. Es kommt in weiten Teilen zu Hybridisation. Einziger erkennbarer Isolationsfaktor ist die Tatsache, dass die major-Vögel überwiegend Standvögel sind und die minor-Vögel Zugvögel" (Bernd Petri, brieflich 2001).