
5) Ist die Variabilität der Frucht-Verbreitungsmechanismen von Scabiosa für die Entstehung der Utricularia-Falle relevant?
Vorbemerkung: In der nachstehend aufgeführten Diskussion habe ich alle wissenschaftliche Hinweise und Argumente der Professoren C. und D. wiedergegeben, die sie 1994 zu meiner Artbegriffsarbeit verfasst hatten, sowie meine Kommentare dazu. Dazu kommen einige Nachträge (N.: Nachtrag; Überschriften 1), 2)…zur Gliederung von mir).
Hier sei noch einmal betont, dass die Artbegriffsarbeit de facto ein 622-Seiten-Exposé zur Frage ist, inwieweit der Species "feste Grenzen gesteckt sind, über welche hinaus sie sich nicht zu ändern vermag".
Im Zusammenhang mit der vorliegenden Mendel-Arbeit möchte ich den Leser wieder bitten, beim Studium der Diskussion die Frage im Sinn zu behalten, wie Gutachter mit einer ausgeprägt darwinistischen Grundhaltung seinerzeit Mendels Arbeit von 1866 beurteilt h a b e n (als sie u. a. 67mal in der kurzen Pisum-Arbeit das Adjektiv "constant" in Verbindungen wie "constant differierende Merkmale", "constante Nachkommen", "constante Formen", "constante Verbindungen, "constante Combinationsformen" etc. gelesen und darüber hinaus festgestellt hatten, dass Mendel in der Entdeckung der Vererbungsgesetze eine Bestätigung von Gärtners Auffassung sah, "dass der Species feste Grenzen gesteckt sind, über welche hinaus sie sich nicht zu ändern vermag"); - und weiter: wie solche Gutachter ein bestätigendes Exposé heute beurteilen würden!
Hat die ausgeprägt darwinistischer Einstellung bedeutender Autoritäten (wie sie in der folgenden Diskussion zum Ausdruck kommt) eventuell zu ähnlichen Missverständnissen geführt wie seinerzeit bei den darwinistischen (und weiteren evolutionistischen) Autoritäten bei der Beurteilung der Mendelschen Befunde? Der Leser urteile bitte wieder selbst!
(Mein Brief vom 26. 8. 1994 an Herrn Prof. D.:)
(W.-E.L.:) "Sehr geehrter Herr D.!
Beiliegend finden Sie meine Diskussion der Einwände von Herrn Prof. C.. Soweit ich das bisher verstehen kann, ist keiner der Punkte wirklich stichhaltig. Aber ich lasse mich aufgrund biologischer Tatsachen auch gern eines Besseren belehren und ich lade Sie und Herrn C. ein, mir zu den hier diskutierten Punkten solche Tatsachen zu nennen.
Sobald ich Zeit dafür finde, möchte ich gerne auch detailliert auf Ihre Kommentare zu sprechen kommen. Vorweg möchte ich jedoch schon einen Punkt erwähnen, der mir besonders am Herzen liegt: Meine Arbeit resultiert nicht aus der aus den USA kommenden Kreationismus-Debatte (zu der man das vorliegende Werk daher auch nicht rechnen kann). Als Student - ich habe von 1965-1971 studiert - habe ich das 1953 erschienene 1300 Seiten umfassende evolutionskritische Werk SYNTHETISCHE ARTBILDUNG von Heribert Nilsson (Genetiker und Professor für Botanik an der Universität Lund in Schweden) so gründlich studiert, dass ich viele Passagen auswendig konnte. Das Werk ist meines Wissens ohne religiöse Zielrichtung und ohne religiösen Hintergrund im großen schwedischen (naturwissenschaftlichen) Verlag Gleerups erschienen und war zuvor von naturwissenschaftlichen Fonds gefördert worden. Das Werk enthält keinerlei Diskussion religiöser Fragen.
Damals war vom amerikanischen Kreationismus in Europa noch nicht die Rede.
Als zweites wissenschaftliches Werk - ebenfalls religiös neutral - habe ich genauso gründlich die Arbeit THE TRANSFORMIST ILLUSION (1957) des englischen Zoologen Douglas Dewar studiert. Ich habe während meiner Studienzeit dann alle möglichen (und unmöglichen) Arbeiten zu dieser Frage gesammelt, wobei vor allem deutschsprachige Autoren mich stärker beeindruckten, so z. B. die in katholischen Verlagen erschienenen, aber doch in religiösen Fragen eher zurückhaltenden, biologischen Arbeiten von Hedwig Conrad-Martius (1947), Oskar Kuhn (1950) und Robert Nachtwey (1959). (Die Schriften sind zum größten Teil in religiösen Fragen so zurückhaltend wie etwa die heutigen biologischen Werke aus dem Herder-Verlag.) Man könnte natürlich einwenden, dass die Abstammungslehre selbst eine religiöse Frage ist. Aber aus dieser Sicht fallen auch alle neodarwinistischen Beiträge unter diese Rubrik.
Alle diese Studien stammen aus 'vor-kreationistischer' Zeit. Als dann später die ersten Traktate und Schriften des Kreationismus in Deutschland publiziert wurden, habe ich auch diese (sowie die amerikanische Literatur) in mein Studienprogramm mit einbezogen - übrigens durchaus kritisch - vgl. Sie bitte das von Ihnen bisher wenig verstandene Unterkapitel S. 271ff. "Der Artbegriff der Schöpfungslehre", in dem ich speziell den Artbegriff des 'amerikanischen Kreationismus' systematisch untersuche und seine Schwächen aufzeige. Weitere kritische Anmerkungen zum Kreationismus finden Sie auf der Seite 53 (Fußnote) und S. 282.
Soviel vorweg. Bedanken möchte ich mich jedenfalls für Ihre Arbeit (acht Seiten Kommentare!), die Sie sich mit meinem Beitrag zum Artbegriff gemacht haben, auch wenn wir vielen Punkten verschiedener Meinung sind."
(W.-E.L.)
(Drei Tage zuvor hatte ich den folgenden Brief zunächst an Herrn Prof. C. geschickt, denn ich konzentriere mich dort auf seine Kommentare. In der Anrede werden jedoch schon beide Herren angesprochen, da von beiden Kommentare gekommen waren. Hier also mein Brief vom 23. 8. 1994 an die Herren C. und D.)
(W.-E.L.:) "Sehr geehrter Herr C.!
Sehr geehrter Herr D.!
Für Ihre Bemühungen, eine Stellungnahme zu meiner Arbeit Artbegriff, Evolution und Schöpfung zu verfassen ( -meines "interessanten und provokativen Buches", wie Herr C. es nennt -) möchte ich mich sehr herzlich bedanken. Es bedarf ja in der Regel eines besonderen Aufwandes an Geist, Kraft und Zeit, eine ausführliche und häufig bis ins Detail naturwissenschaftlich begründete Auffassung zum Ursprung der Lebensformen, mit der man in ganz wesentlichen Punkten nicht übereinstimmt, zu studieren und differenziert zu beurteilen.
Da ich nun meinerseits manchen Punkten Ihrer Beurteilung (aus guten naturwissenschaftlichen und logischen Gründen, behaupte ich vorweg) nicht folgen kann und Sie vorgeschlagen haben, dass "die in unseren Stellungnahmen enthaltenen Hinweise...tatsächlich berücksichtigt und integriert werden", möchte ich diesem Vorschlag gerne folgen und mir erlauben, Ihren Text zunächst wörtlich zu zitieren und anschließend zu diskutieren.
(Prof. C.:) "Nun aber zu Lönnig's Buch "Artbegriff, Evolution und Schöpfung" (3. Auflage 1988). Dazu hat Herr Prof. D. (an unserem Institut) eine sehr ausführliche Stellungnahme ausgearbeitet, der ich mich vollinhaltlich anschließen möchte. Prof. D. ist einer meiner ersten Dissertanten und hat im Rahmen seines Doktoratsstudiums das Nebenfach "Philosophie" absolviert. Er ist also im Hinblick auf eine Beurteilung des interessanten und provokativen Buches noch besser qualifiziert als ich selbst. Trotzdem muss auch er einräumen, dass eine umfassende und allen Aspekten gerechtwerdende Beurteilung nur aus einem Gutachterteam kommen könnte, dem außer einem Evolutionsbiologen auch ein einschlägig interessierter Philosoph und ein Theologe angehören müssten. "
(W.-E.L.:) Wenn wirklich so viele Koryphäen notwendig sein sollten, um das Buch zu beurteilen, könnte ich das vielleicht als besonderes Kompliment für die Bandbreite und Qualität meiner Arbeit auf vielen Gebieten auffassen. Aber ich bin nicht der Auffassung, dass es erst eines einschlägig interessierten Philosophen und eines Theologen bedarf, um die in allen wesentlichen naturwissenschaftlichen Begründungen und weiteren Schlussfolgerungen denkbar klare und für Biologen zumeist einfache Arbeit zu beurteilen.
Da die Frage nach einem Gutachten von den Professoren E. und F. zunächst im Zusammenhang mit einer eventuellen Habilitation* im Fach Genetik an der Universität zu Köln vorgeschlagen worden ist, verstehe ich Ihre Meinung als ablehnende Antwort auf diese Frage; denn welcher Kölner Genetiker würde mit einem Philosophen und Theologen zusammen ein Gutachten zur Habilitation an der hiesigen naturwissenschaftlichen Fakultät verfassen ? !
(*N.: Wegen der herrschenden neodarwinistischen Auffassung bin ich auf den Gedanken, diese Arbeit als Habilitations-Arbeit vorzuschlagen, selbst nicht gekommen. Ich habe die Idee jedoch aufgegriffen und weiterverfolgt, nachdem Prof. E. diesen Vorschlag gemacht hatte. Weiter sollte ich vielleicht noch erwähnen, dass sich auch relativ schnell zwei C4-Professoren für positive Gutachten - im Sinne Waesberghes siehe unten - fanden. Darüber hinaus gibt es noch zwei unabhängige positive Gutachten, die ein wissenschaftlicher Verlag eingeholt hat.)
(Prof. C.:) "Hier noch einige kurze persönliche Bemerkungen: Als Evolutionsbiologe kann man sicherlich die Intensionen von Lönnig am besten nachvollziehen, wenn man seine Diskussion der Kommentare und Einwände von PD Dr. S.L. auf den Seiten 588 ff. studiert. Hier wird immer wieder behauptet, dass Arten mit "Gewinn an genetischer Information, mit Neukonstruktionen samt Synorganisationen auf den verschiedensten Ebenen" nur "durch einen intelligenten Schöpfungsakt" entstehen können. Zu diesem Thema haben wir in den letzten Jahren zusammen mit Frau Dr. V. Mayer die stammesgeschichtliche Entwicklung von einfachen, abfallenden Schließfrüchten zu hochspezialisierten Fallschirmfliegern bei Arten der Gattung Scabiosa (Dipsacaceae) mit multidisziplinären Methoden und Experimenten analysiert. Dabei kann die primäre Ausbildung von Außenkelchsäumen als Zusatzassimilatoren für die heranwachsenden Früchte verstanden werden, die erst ab einer gewissen Größe im Sinne eines Funktionswandels auch als fallgeschwindigkeitsverzögernde Flugorgane für die reifen Früchte in Erscheinung treten. Wenn wir nur die Anfangs- und die Endglieder dieser Entwicklungsreihe zur Verfügung hätten, niemand würde den durch viele kleine Veränderungsschritte in verschiedenen Bereichen erfolgten Übergang zur synorganisierten Neukonstruktion für möglich halten. Ähnlich liegen offenbar auch die Fälle bei komplexen und hochspezialisierten Organen, wie etwa den Tierfallen bei Utricularia oder Nepenthes."
(W.-E.L.:) Für die Xerokopie der Dissertation von Frau V. Mayer möchte ich mich an dieser Stelle noch einmal herzlich bedanken. Ein paar Gedanken zu Ihren Hauptthesen:
1) Entwicklung durch viele kleine Schritte von einfachen, abfallenden Schließfrüchten zu hochspezialisierten Fallschirmfliegern bei Arten der Gattung Scabiosa;
2)Außenkelchsäume: zunächst Zusatzassimilatoren, dann Funktionswandel zu Flugorganen
.Auf der nächsten Seite habe ich alle 17 Abbildungen von Scabiosa Früchten aus der Dissertation von Frau Mayer zusammen wiedergegeben, so dass man die dort dargestellte Variabilität mit einem Blick erfassen kann. Was mir zunächst auffällt, ist das von Troll gern zitierte "Prinzip der variablen Proportionen" innerhalb eines Bauplans, nicht aber eine Entwicklung über viele kleine Schritte von vielleicht einem Bauplan oder Typus zu einem ganz anderen. Alle Scabiosa-Früchte sind z. B. durch einen Außenkelch plus Schirm gekennzeichnet. Variieren kann nun die Frucht-, Schirm und Grannengröße, die Epidermisstruktur und viele weitere Parameter (wie das Frau Mayer bis ins Detail in ihrer gründlichen Studie aufgezeigt hat - von der Zahl und Verteilung der Stomata bis zu den Sclerenchymelementen, Aufbau des Perikarps etc.). Der Typus (Troll) bleibt davon praktisch unberührt. Darüber hinaus weist Frau Mayer in ihrer Arbeit darauf hin, dass sich (innerhalb dieses Formenkreises) die unterschiedliche Gestaltung des Außenkelchs der verschiedenen Sectionen von Scabiosa ohne Übergangsformen darstellt (S.87).
Einige Details zur Frage des Funktionswandels: "Keine Bedeutung für die Anemochorie haben dagegen die schmalen Schirme bei Früchten der sect. Scabiosa " (V. Mayer 1993, S. 88 und folgende Argumentation mit Tabelle 12).
Abb. 5
Zusammenstellung aller in V. Mayers Dissertation aufgeführten Abbildungen von Scabiosa-Früchten (Abb. 1: 4 a,b,c, und Abb. 8 und 9; Namen siehe dort).
Ich halte diese Aussage für möglich und will das auch der Argumentation halber vorerst einmal annehmen (obwohl ich bei dieser Interpretation Schwierigkeiten mit den Tabellen 15 und 16 habe, in denen die Arten der sect. Scabiosa - S. trianda, S. cinerea und S. ochroleuca (S. 61) und S. cinerea und S. ochroleuca (S. 62) -zu den 6 (von 15) bzw. 3 (von 11) Scabiosa-Arten mit der weitesten Ausbreitungsdistanz gehören und damit großschirmige Arten wie S. stellata, S. caucasica und andere zum Teil sogar weit übertreffen; - darüber hinaus stellt sich vielleicht auch die Frage, ob nicht kleinschirmige Formen auf dem Boden ebenfalls besser vom Wind erfasst werden können als schirmlose Samen).
Wie dem auch sei - nach Herrn C.s und Frau Mayers Interpretation (S. 88) "wäre dann auch die Vergrößerung des häutigen Schirmes primär auf eine Oberflächenvergrößerung für die Photosynthese zurückzuführen" (S. 88/89). Das halte ich aufgrund folgender Tatsachen für unwahrscheinlich: "Während bei S. rotata und S. prolifera...die CO2-Fixierungsrate der Köpfchen mit Involucralblättern bei 3 -16 liegt, wird bei S. cinerea und S. ochroleuca auch bei jüngeren Pflanzen mit einer relativ kleinen Rosette 99,3 bis 99,7% der Assimilation von den Laubblättern übernommen (Tab. 10)" (Mayer 1993, S. 43 ). Ohne Involucrum reduziert sich dann der Anteil des Schirms noch einmal ganz erheblich (wobei man noch zusätzlich zwischen Außenkelchtubus und Schirm differenzieren könnte oder sogar müsste; denn wenn man mit einem für die postulierte Selektion relevanten Photosynthesebeitrag der 2-schichtigen, häutigen Gewebe der Rippenzwischenräume rechnet (S.88), dann muss man wohl den chloroplastenhaltigen Tubus bei der Gesamtbilanz ebenfalls berücksichtigen).
Der entscheidende Punkt ist nun, dass ein eindeutiger, eine komplexe Entwicklungsrichtung bestimmender Selektionsvorteil mit weniger als 0.3 bis 0,7% CO2-Assimilierungsrate selbst bei schon deutlich ausgebildeten und bis ins Detail differenzierten Schirmen (wie bei S. cinerea und S. ochroleuca festzustellen) nun weder bewiesen noch sinnvoll nachvollzogen werden kann. Wenn es tatsächlich darum ginge, die CO2-Assimilierung sagen wir um 1% für die ganze Pflanze zu erhöhen, dann ist das sehr viel einfacher mit einigen Mitosen in den Blattspreiten oder der stärkeren Konzentration von Chloroplasten in den Stengeln oder in den Involucralblättern etc, zu bewerkstelligen. Dazu bedarf es doch nicht des Aufbaus einer neuen Struktur (oder eines neues Systems von Strukturen), hier des Schirmes, an dessen Bildung vermutlich mehrere in Sequenz und Funktion aufeinander abgestimmte Gene beteiligt sind (für das Gesamtsystem von Außen- und Innenkelch vermutlich auch einige völlig neue Struktur- und/oder Regulatorsequenzen).
Rechnet man nun für die Entwicklung des Schirms noch mit "vielen kleinen Schritten", mit "slight or even invisible effects on the phenotype" (Mayr), also mit einer kontinuierlichen Entwicklung, mit fließenden Übergängen, dann müsste der Beitrag zur Assimilation anfangs noch sehr viel geringer gewesen sein als bei S. cinerea und S. ochroleuca mit ihren schon relativ gut ausgebildeten Schirmen. Welchen Selektionsvorteil sollen aber Beiträge von 0,00 1% oder 0,0015% oder 0.003% etc. zur Assimilationsrate der ganze Pflanze und der jeweiligen Spezies von winzigsten Schirmanfängen bis zur sect. Scabiosa gehabt haben? Die Variabilität der Assimilationsleistungen (ohne Schirm oder Schirmanfänge) dürfte schon innerhalb einer Spezies aufgrund unterschiedlichen Pflanzenwuchses an den verschiedensten Standorten solche anfänglichen "Schirmbeiträge" um ein Vielfaches übertroffen haben. In einer solchen Kosten-Nutzen-Rechnung müssten weiter die Energiekosten für den Schirmaufbau eingehen, die aufgrund der Schirmkomplexität sehr wahrscheinlich höher sind als bei einigen zusätzlich Mitosen und/oder höheren Chloroplastenkonzentrationen in schon bestehenden assimilatorischen Geweben. Es spricht daher meiner Auffassung nach wesentlich mehr gegen eine solche Entwicklung in vielen kleinen Schritten (primär als eine Oberflächenvergrößerung für die Photosynthese - wobei für jeden kleinen Schritt deutliche Selektionsvorteile gefordert werden müssten) als dafür.
Die hier aufgeführten selektionistischen Schwierigkeiten für die Bildung der Schirme lassen sich im Prinzip auch auf die Entstehung des gesamten Außenkelchs der Dipsacaceen übertragen, zumal 'die Spaltöffnungen frühestens zum Zeitpunkt der Blüte beobachtbar und nicht über den gesamten Epicalyx verteilt sind, wie bei einer aus Hochblättern entstandenen Struktur durchaus zu erwarten gewesen wäre' (Mayer 1993, S. 28). Das würde ja sogar bedeuten, dass ein Teil der Photosyntheseaktivitäten bei der Entwicklung des Außenkelchs erst einmal aufgegeben und darüber hinaus dieser Verlust sogar bis heute beibehalten wurde, was die Selektionshypothese von der photosynthetischen Oberflächenvergrößerung in kontinuierlicher Entwicklung (d. h. in zahlreichen kleinen Schritten) zu den Fallschirmstrukturen von Scabiosa noch unwahrscheinlicher macht.
Es dürfte auch schwierig sein, einen kontinuierlich ablaufenden Evolutionsprozess von vier Hochblättern zum Außenkelch der Dipsacaceen selektionistisch überzeugend zu begründen; denn wieder müsste jeder der vielen kleinen Schritte der sich aufeinander und auf die Kelchblätter zu bewegenden Hochblätter einen deutlichen Selektionsvorteil gegenüber den jeweiligen Vorstufen gehabt haben. Dagegen könnte z.B. die schrittweise zunehmende Beschattung das angenommenen zweiten (unteren) Hochblattpaars (Verlust an photosynthetisch optimal arbeitender Blattfläche) angeführt werden.
Völlig unbekannt sind bisher die molekularbiologischen Grundlagen für die Außen- und Innenkelchstrukturen der Dipsacaceen. Wie schon oben erwähnt, dürfte es sich - etwa in Analogie zum Anthocyansystem - um ein komplexes Gefüge aufeinander abgestimmter DNA-Sequenzen und -Funktionen handeln, wobei auch einige völlig neue Sequenzen im Vergleich zu anderen Pflanzenfamilien denkbar sind. In einem solchen System von Regulator- und Targetgenen kann nun Allelbildung zu zahlreichen Abwandlungen in der Schirmgröße, Stomataverteilung, Sklerenchymbildung etc. führen -und das jeweils in mehr oder minder 'großen' (Mutations-) Schritten und Kombinationen von Allelen mit in der Regel deutlich sichtbaren Auswirkungen auf den Phänotyp. Dabei brauchen die verschiedenen Allele keinesfalls alle einen positiven Selektionswert haben, wie das die Neutralitätstheorie überzeugend aufgezeigt hat. Nachweislich können sich "slightly deleterious genes" und sogar der Ausfall von Genfunktionen en masse 'behaupten' (vgl. Artbegriffsarbeit z.B. S. 193, 403-419, 423). Nach einer (durch einen intelligenten Schöpfungsakt) erfolgten Neuorganisation des für den Typus der Dipsacaceen grundlegenden genetischen Sequenz- und Funktionsgefüges war das Potential für zahlreiche Abwandlungen innerhalb der Familie und der Gattungen geschaffen worden. Gen-, Chromosomen und Genommutationen, Auto- und Allopolyploidie, Transposons und (wohl weniger häufig) Genduplikationen, Heterosis und sekundäre Sterilitätsbarrieren konnten (mit und ohne Selektionsvorteile) zahlreiche Formen der Degeneration und Anpassung bewerkstelligen.
(Im übrigen ist auch innerhalb der Familie der Dipsacaceen keine Entwicklung durch viele kleine Veränderungsschritte nachweisbar. Vielmehr scheinen große Schritte durch (Regulator-)Gen-, Chromosomen und Genommutationen sowie Allopolyploide die entscheidende Rolle gespielt zu haben; dennoch bleiben auch innerhalb der Familie noch viele Fragen offen, so dass Herr Prof. C. zu dieser Frage zwar von den "vermutlichen phylogenetischen Zusammenhängen zwischen den wichtigsten Sippengruppen" (1991, S. 518, Lehrbuch) sprechen, aber kein Kontinuum nachweisen kann.)
In das umfangreiche Tatsachenmaterial zur Degeneration der Organismen lassen sich nun auch mühelos die mehr oder minder (oder vielleicht auch völlig) funktionslosen Schirme der Außenkelche von Scabiosa, sect. Scabiosa, integrieren. Die oft bis ins Detail fragwürdigen Selektionsthesen einer Entwicklung über viele kleine Schritte können wir biologisch gut begründet fallen lassen.
Fazit: Die als kontinuierliche Evolution (durch viele kleine Veränderungsschritte) postulierte "stammesgeschichtliche Entwicklung von einfachen, abfallenden Schließfrüchten zu hochspezialisierten Fallschirmfliegern der Arten der Gattung Scabiosa (Dipsacaceae)" hat vermutlich niemals stattgefunden. Aufgrund zahlreicher Beispiele für Degenerationsprozesse im Organismenreich (darunter viele Fälle von Organisations- und Komplexitätsverlusten von Samenverbreitungsstrukturen z. B. auf ozeanischen Inseln) und des Defektcharakters der meisten Mutationen ist es hingegen denkbar, dass der Prozess umgekehrt verlaufen ist: Von der synorganisierten Neukonstruktion hochspezialisierter Fallschirmflieger zu einfachen abfallenden Schließfrüchten. Nach der obigen Analyse jedenfalls "kann die primäre Ausbildung von Außenkelchsäumen nicht a 1 s Zusatzassimilatoren für die heranwachsenden Früchte verstanden werden, die erst ab einer gewissen Größe im Sinne eines Funktionswechsels auch als fallgeschwindigkeitsverzögernde Flugorgane für die reifen Früchte in Erscheinung treten". Die These: "Wenn wir nur die Anfangs und Endglieder dieser Entwicklungsreihe zur Verfügung hätten, niemand würde den durch viele kleine Veränderungsschritte in vielen Bereichen erfolgten Übergang zur synorganisierten Neukonstruktion für möglich halten" entbehrt nach meinem bisherigen Verständnis der faktisch-biologischen Grundlage: Weder ist eine Entwicklung durch viele kleine Veränderungsschritte innerhalb der Gattung Scabiosa nachgewiesen (der Außenkelch der verschiedenen Sektionen von Scabiosa präsentiert sich "ohne Übergangsformen" Mayer 1993, S. 86), noch hat ein grundlegender Bauplanwechsel stattgefunden. Darüber hinaus könnte man sich eine schrittweise Vergrößerung des Schirms und Verlängerung der Grannen des Innenkelches etc. ohne weiteres "vorstellen" (ob diese Vorstellungen dann mit der Realität kongruent wären, ist eine andere Frage) und die Schlussfolgerung, dass ohne Kenntnis der "Anfangs und Endglieder" niemand den "Übergang zur synorganisierten Neukonstruktion für möglich halten würde" erscheint mir daher auch nicht überzeugend.
"Ähnlich liegen offenbar auch die Fälle bei komplexen und hochspezialisierten Organen, wie etwa den Tierfallen bei Utricularia oder Nepenthes." - Abgesehen davon, dass es sich bei der stammesgeschichtlichen Entwicklung in zahlreichen kleinen Schritten des Scabiosa-Beispiels um eine neodarwinistische Hypothese handelt, die in den Hauptpunkten nicht durch biologische Tatsachen abgesichert werden kann (kein Kontinuum, Fragwürdigkeit der positiven Selektionswerte postulierter Anfangs- und Übergangsstadien, Fragwürdigkeit des Funktionswechsels), sind die Fälle Utricularia und Nepenthes wesentlich komplexer! Bei Utricularia ist nun tatsächlich ein totaler Bauplanwechsel zu konstatieren. Überdies ist das Funktionsgefüge der Tierfallen hier so komplex, dass es - im Gegensatz zu den 'Fallschirmen' von Scabiosa -bis heute nicht völlig verstanden und erklärt werden kann. Weiter sind die Strukturen innerhalb des Funktionsgefüges der Falle derart verkoppelt, dass zahlreiche Biologen - auch Vertreter der Synthetischen Evolutionstheorie - im Prinzip immer wieder folgendes zu Utricularia bekannt haben:"...no adequate evolutionary sequence can yet be constructed even to present a speculative path for the origin of what appears to be a relatively homogeneous group" (Juniper/Robins/Joel: Carnivorous Plants; 1989, p. 43). Wie schon erwähnt, kann man sich ganz im Gegensatz zu Utricularia allein schon bei Kenntnis der angenommenen Anfangs- und Endformen von Außenkelchsäumen und Innenkelchstrukturen bei Scabiosa sehr wohl einen "speculative path" vorstellen, wie sich eine angenommene Entwicklung in vielen kleinen Schritten abgespielt haben könnte (auch wenn bei genauerer Betrachtung die positiven Selektionswerte der kleinen Schritte fragwürdig sind).
Man würde also besser sagen können: "Hinsichtlich des Komplexitätsgrades und der "Vorstellungsmöglichkeiten" völlig anders als innerhalb der Gattung Scabiosa liegen die Fälle bei komplexen und hochspezialisierten Organen, wie etwa bei den Tierfallen bei Utricularia oder Nepenthes. "
Im folgenden einige Daten und Fragen zu Utricularia von R. Nachtwey. Soweit ich informiert bin, hat bislang niemand seine Einwände im Sinne der Synthetischen Evolutionstheorie befriedigend beantworten können.
(N.: Zur einfachsten Handhabung für den Leser sei hier Abbildung und Text wiederholt, wie oben, aber kleiner:)
Abb. 6
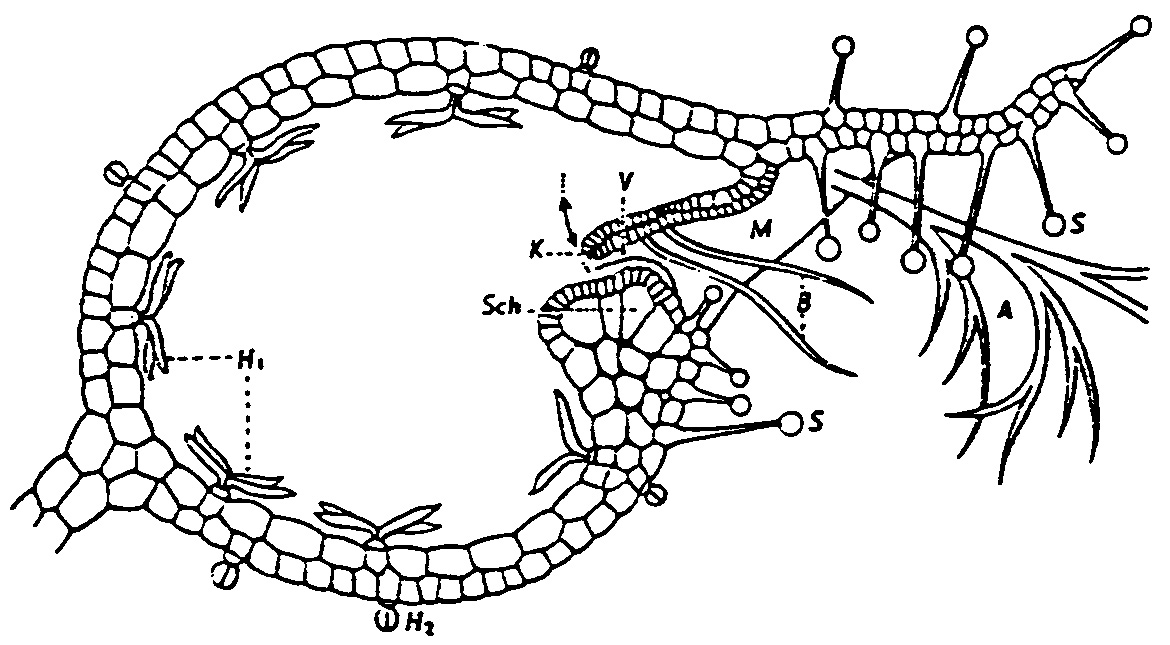
Abb..95. Utricularia. Längsschnitt durch die Blase. Schematisiert, zum Teil nach LLOYD. Klappe (zur Verdeutlichung) etwas angehoben gezeichnet; Gefäßbindel nicht gezeichnet. M Mündung bzw. Blaseneingang; K Verschlussklappe; V abdichtendes Velum; Sch Schwelle (Widerlager); B Borsten; .A Antenne; H1 vierarmige Haare; H2 Drüsenköpfchen.
(
Aus: Th. Schmucker und G. Linnemann 1959, p. 251)"In den Moorgräben wächst der Wasserschlauch (Utricularia vulgaris L.), der mit Hilfe äußerst sinnreich konstruierter Bläschen, die als winzige Kastenfallen funktionieren, massenhaft kleine Wassertiere fängt und sie verdaut. Tausende solcher Bläschen sitzen an einer einzigen Pflanze. Die kleine Tierfalle misst zwei bis vier Millimeter...Die Eingangsöffnung des fast eiförmig gestalteten Bläschens ist durch eine Klappe verschlossen. Diese Klapptür ruht unten auf einem hufeisenförmigen Widerlager und kann sich nur nach innen öffnen. An ihrer Außenwand trägt die Klappe keulenförmige Drüsenhaare und reizbare, schlanke Sinneshaare. Flankiert wird sie jederseits von einem grünen verzweigten Ästchen. Das Bläschen ist vor dem Beutefang stets in einer gespannten Fangstellung, seine Seitenwände sind dann weit nach innen gewölbt, und das Innere ist von Wasser leergepumpt. Die Wände sind nämlich von einer Wasserleitung durchzogen und imstande, in etwa einer Viertelstunde die Wasserfüllung des Bläschens völlig aufzusaugen.
Die vom Wasserschlauch bewohnten Moorgewässer sind von Kleinkrebsen in großer Zahl bevölkert. Winzige Hüpferlinge (Copepoden), aber auch Zweighornkrebschen (Cladoceren) und Muschelkrebschen (Ostracoden) tummeln sich zwischen den schwimmenden Wasserschlauchpflanzen, die eine Länge von über einem halben Meter erreichen. Die keulenförmigen Drüsenhaare der Bläschenklappe sondern einen Schleim ab, dessen Bedeutung noch unbekannt ist, der aber vielleicht für die Krebschen etwas Anziehendes hat. Sobald ein Krebschen eines der schlanken Sinneshaare berührt, ist es schon verloren. Mit einem blitzschnellen Ruck bewegen sich die unter beträchtlicher Spannung stehenden Blasenwände nach außen. Die Klappe schnellt nach innen. Das vor dem Blaseneingang befindliche Wasser wird samt dem Beutetierchen in das Blaseninnere gerissen, und sogleich schließt sich die Klapptür wieder. Der ganze Vorgang spielt sich in einem Zeitraum von 1/100 bis 1/200 Sekunde ab (nach neuesten Erkenntnisse 1/500 Sekunde; Anmerkung W.-E.L.). Der Sog ist so heftig, dass selbst Insektenlarven und Würmer, deren Länge die des Bläschens mehrmals übertrifft, in die Kastenfalle hineingerissen werden. Im Teufelsmoor bei Bremen fand ich im Wasserschlauchbläschen eng zusammengeringelte Borstenwürmer, die das Bläschen durch ihre Größe unförmig auftrieben. Sie waren schon teilweise verdaut. Auch diese abnorm großen Beutetiere werden durch den plötzlichen, gewaltigen Sog gefangen. Die entscheidende Kraft des Soges scheint von den seitlichen Blasenwänden auszugehen. Wie ihre schnelle Reaktion möglich ist, konnte bisher nicht geklärt werden.
Bevor der Verdauungsvorgang einsetzt, saugt die Pflanze wieder das gesamte Wasser aus dem gefüllten Bläschen heraus. Die Klapptür schließt so dicht, dass von außen kein neues Wasser eindringt. Die Verdauungssäfte können also unverdünnt auf die Beute einwirken. Sie werden jetzt von eigenartigen, vierstrahlig angeordneten Drüsenschläuchen ausgegossen, die der inneren Wandfläche massenhaft aufsitzen. Ein eiweißlösendes Ferment und Benzoesäure werden von den Drüsen erzeugt, wie von Lützelburg zuerst nachgewiesen hat, und so kann das Bläschen nun wie ein richtiger tierischer Magen arbeiten und die Beute regelrecht verdauen. Die Benzoesäure, die bei der Verdauung wie die Salzsäure unseres Magens mitwirkt, erfüllt auch den Zweck, Fäulnis im Bläschen zu verhüten. Das Fleisch der Opfer wird in ganz kurzer Zeit verdaut, die Eiweißbausteine der Krebschen werden aufgesogen und vom Wasserschlauch zu arteigenem, pflanzlichen Eiweiß wieder zusammengebaut. Die mit Krebschen ernährten Wasserschlauchpflanzen gedeihen weit besser als die, denen solche Nahrung entzogen wird. Stutzer gelang es auch, drei Arten von Colibazillen in den Bläschen aufzufinden. Das ist ein besonderer Beweis dafür, dass die Funktion der Bläschen der Wirkungsweise eines tierischen Darmkanals hochgradig ähnlich ist. Es handelt sich nämlich um genau dieselben Colibakterien, die im Darm von Tieren und Menschen beständig anwesend sind. Sie verhindern die Vermehrung fäulniserregender Bakterien. Die Wasserschlauchpflanze hat sich also Tausende von kleinen tierischen Mägen angeschafft.
Ist der Verdauungs- und Aufsaugungsvorgang beendet, so kehrt das Bläschen wieder in seine hochgespannte Fangstellung zurück und erwartet die nächste Beute. Die unverdaulichen Panzer der Krebschen bleiben im Bläschen. Erst wenn dieses fast ganz mit harten Resten gefüllt ist, fällt es ab. Fast stets sind in einem Bläschen mehrere Opfer zu finden, ja sogar häufig 10 -20 Krebschen, die jedoch alle einzeln gefangen wurden. Es hat also ein totaler Funktionswechsel stattgefunden. Ein normales Blatt, Organ der pflanzlichen Assimilation, das die Umwandlung anorganischer Stoffe in organische mit Hilfe des Lichtes ausführt, hat einen kleinen Fangapparat mit allen mechanischen Einrichtungen gebildet, der zugleich wie ein tierischer Magen arbeiten kann und alle dafür notwendigen Säfte besitzt.
Wie wir im vorigen Kapitel gesehen haben, lehrt der moderne Darwinismus, dass alle organischen Bildungen durch "Mikroevolution" entstanden seien. Dabei wird ausdrücklich betont, dass winzigste Mutationen das Material für die natürliche Selektion liefern. Ludwig betont, dass die Kleinstmutationen ohne oder fast ohne erkennbaren Effekt sind. "Unter diesen Kleinstmutationen wird man den Hauptteil der evolutorisch verwertbaren Mutationen zu suchen haben, und nur sehr selten sind günstige Großmutationen zu erwarten."
Nun mögen uns die Darwinisten erklären, wie man sich die Bildung des Wasserschlauchbläschens aus einem Blattzipfel vorstellen soll. Welche richtungslose Mutation soll im normalen Blattzipfel zuerst erfolgt sein und dann irgendeinen Auslesewert gehabt haben? Hatte sie diesen nicht, so ging sie als belanglos verloren. Ausdrücklich betonen die Darwinisten, dass Mutation und Selektion zusammenwirken müssen, wenn etwas Neues entstehen soll. Durch welche blind zusammengewürfelten, richtungslosen Kleinmutationen soll das Wasserschlauchbläschen entstanden sein?...Wie sollen wir uns den ersten Anfang zur Bildung dieser Kastenfalle, die wie ein tierischer Magen arbeitet, eigentlich vorstellen?...
Soll die Bildung mit dem Entstehen der Kastenfalle beginnen oder mit der Produktion der Verdauungssäfte? - Sobald wir dies überlegen, zeigt sich die...Ohnmacht der Darwinschen Theorie, denn selbst eine vollkommene Kastenfalle mit der erstaunlichsten Fähigkeit, blitzschnell Tiere zu erbeuten, hätte ohne Verdauungssäfte nicht den geringsten Wert im Daseinskampf, weil die Beute nicht verdaut würde. Was aber soll es andererseits einem gewöhnlichen Blattzipfel nützen, wenn er noch so wirksame Verdauungssäfte ausscheidet, er kann ja die Beute nicht festhalten, was unbedingt nötig ist. Aber selbst wenn Kastenfalle und Verdauungssäfte zusammenwirken, so ist für den Daseinskampf noch nichts gewonnen...Die gelösten Eiweißstoffe müssen ja auch aufgesogen und in arteigenes Pflanzeneiweiß verwandelt werden…Die Bildung des Wasserschlauchbläschens erfordert also das vollendet harmonische Zusammenspiel vieler verschiedenartiger Gene und Entwicklungsfaktoren. Erst mit dem Endeffekt wird der Nutzen für den Daseinskampf erreicht, nicht aber mit irgendeiner Entwicklungsstufe" (von Nachtwey kursiv).
Ich möchte abschließend hervorheben, dass mit dem Hinweis auf die Verhältnisse von Scabiosa selbst dann keine befriedigende Antwort auf die Problematik bei Utricularia gegeben wäre, wenn Scabiosa nachweislich (empirisch) und vollständig im neodarwinistischen Sinne erklärt werden könnte. Der Komplexitätsgrad liegt bei Utricularia wesentlich höher und verlangt eine eigene Untersuchung und Bewertung.
Selbstverständlich würde mich auch interessieren, wie Sie Nachtweys oben zitierte Fragen (zunächst einmal) theoretisch beantworten.