Zurück zur Homepage
Die Mendelschen Gesetze in Erwin Baurs EINFÜHRUNG IN DIE
VERERBUNGSLEHRE von
1911: Eine
der frühesten und zugleich besten Darstellungen der Thematik
Baur, E. (1911): Einführung in die
Vererbungslehre. Vorlesung IV. Verlag
von
Gebrüder
Bornträger [damals Berlin W 35, Schöneberger Ufer
12a] (1911, pp. 45-64).
Vorbemerkung von W.-E. Lönnig, Mai 2002: Die folgende Abhandlung der Mendelschen
Gesetze kann als eine der besten
und anschaulichsten 'Vorlesungen' zum Thema überhaupt gelten. Auf einige nomenklatorische Unterschiede
möchte ich
den Leser jedoch aufmerksam machen: Wir sprechen heutzutage nicht mehr allgemein von den Mendelschen
"Spaltungsgesetzen",
sondern differenzieren wie folgt: 1. Für die F1 sprechen wir vom Uniformitätsgesetz
(oder von der Uniformitätsregel), 2. für die F2 vom
Spaltungsgesetz (oder von der Spaltungsregel) und 3. bezeichnen wir das Phänomen von der freien
Kombinierbarkeit
der Gene bei Mehrfaktorenkreuzungen als Unabhängigkeitsregel oder Regel von der Neukombination der
Erbfaktoren (Ausnahmen bei
enger Genkopplung). Statt "homozygotisch" und "heterozygotisch" sagen wir heute homozygot und
heterozygot [Hinweise in eckigen Klammern im Text].
Bei der Dominanzregel schießt Baurs Kritik etwas über
das Ziel hinaus; wir sprechen nach
wie vor von der Dominanzregel und diese beschreibt die große Mehrheit der Fälle: Richtig bleibt
jedoch Baurs Hinweis, dass wir mit genauen Analysemethoden die
heterozygoten von den homozygoten Phänotypen mit dem (oder den) dominanten Merkmal(en) in der
Regel unterscheiden können.
Nun zum Text von Erwin Baur (Baurs Sperrungen sind hier bold und kursiv
wiedergegeben, nur bold und kursiv ist von Baur selbst):
VORLESUNG IV
Die Mendelschen Spaltungsgesetze. - Beispiele von Kreuzung zweier Rassen, die sich in einem Merkmal unterscheiden. - Die Sachlage bei mehrern Unterschieden.
Seit den grundlegenden Versuchen von JOHANNSEN
(189) mit Bohnen- und Gerstenrassen wissen wir, daß auch scheinbar ganz geschlossene systematische Einheiten durchaus nicht einheitlich sind, sondern zusammengesetzt aus einer großen Zahl kleiner Einheiten, den reinen Linien.
........
Wir haben nun schon gehört, daß die Bohnen, an denen JOHANNSEN die kleinsten elementaren Einheiten erkennen konnte, sich durchweg durch Selbstbefruchtung, oder wie der Fachaudruck lautet, "autogam" fortpflanzen. Eine Fremdbefruchtung und damit eine Kreuzung zweier Linien kommt äußerst selten vor. Die reinen Linien bestehen also ganz unvermischt dauernd nebeneinander.
Ein solches dauerndes Isoliertbleiben der einzelnen Linien finden wir außer bei den autogamen auch bei allen sich ausschließlich ungeschlechtlich fortpflanzenden Organismen, z. B. bei den Bakterien, den blaugrünen Algen, vielen völlig apogamen Pflanzengattungen usw. Hier kann man überall eine reine Linie als erblich absolut einheitliches Material sich einfach dadurch verschaffen, daß man ein einzelnes Individuum isoliert und sich vermehren läßt.
So einfach und klar liegen nun aber die Verhältnisse bei den wenigsten Pflanzen und Tieren. Eine große Anzahl von Organismen, vor allem alle getrennt geschlechtlichen, sind auf Fremdbefruchtung angewiesen, sie pflanzen sich "allogam" fort. Die Frage ist da, wie verhält es sich mit der Vererbung bei Fremdbefruchtung, wenn sich Individuen befruchten, die zweierlei verschiedenen systematischen Einheiten angehören? Diese Frage soll uns in der heutigen Vorlesung beschäftigen.
Den Weg für ein tieferes Eindringen in das bis dahin ganz unzugängliche Gebiet der
Vererbung bei Fremdbefruchtung haben die grundlegenden Entdeckungen des Augustinermönches GREGOR MENDEL
(241) gebahnt. Das Verdienst, die ganz in Vergessenheit[?] geratenen von MENDEL gefundenen Gesetze
selbständig
und unabhängig voneinander neu entdeckt zu haben, gebührt den Botanikern C. CORRENS (64), E. TSCHERMAK (374) und H. DE VRIES (387, 388) [genaue historische Reihenfolge: de Vries, Correns, Tschermak].
Um uns mit den von MENDEL gefundenen Vererbungsgesetzen bei Kreuzbefruchtung vertraut zu machen, wollen wir ausgehen von einem Versuch, den jeder selbst leicht wiederholen kann, der über einen kleinen Garten verfügt.

Wir kreuzen zwei Individuen von Antirrhinum majus, dem Gartenlöwenmaul, und zwar ein elfenbeinfarbiges aus einer konstant elfenbeinfarbigen Rasse (Taf. II Fig. 1) und ein rotes aus einer konstant roten Rasse (Taf. II Fig. 2). Für diese beiden Individuen wollen wir eine bestimmte Bezeichnung mit Buchstaben einführen und zwar folgendermaßen: Jede Pflanze entsteht als das Produkt der Vereinigung zweier Sexualzellen, einer männlichen und einer weiblichen. Eine solche Sexualzelle der konstant roten Rasse wollen wir mit F und die durch die Vereinigung zweier solcher Sexualzellen entstandene rote Pflanze mit FF bezeichnen. Ganz entsprechend soll die elfenbeinfarbige Pflanze ff und ihre Sexualzellen f heißen. Wenn wir nun einen Bastard zwischen einem roten Individuum FF und einem elfenbeinfarbigen ff erzeugen, so geschieht das dadurch, daß wir etwa eine weibliche Sexualzelle F sich vereinigen lassen mit einer männlichen f, also in der Weise, daß wir eine rote Pflanze befruchten mit Pollen einer elfenbeinfarbigen. Die Kreuzung kann, das ist ganz einerlei, auch umgekehrt vorgenommen werden. Jedenfalls erhalten wir auf diese Weise ein Individuum mit der Bezeichnung Ff bezw. fF, d. h. einen Bastard oder, wie der Terminus heißt, ein heterozygotisches [heterozygotes] (durch Vereinigung ungleichartiger Sexualzellen entstandenes) Individuum. Im Gegensatz dazu heißt man ein Individuum, das durch die Vereinigung zweier gleichartiger Sexualzellen entstanden ist, homozygotisch [homozygot] . Ein solcher Antirrhinum-Bastard wird nun weder elfenbeinfarbig wie der eine Elter, noch rot wie der andere, sondern blaßrot (Taf. II, Fig. 3) blühen, er hat nur von dem einen - dem roten - Elter her die "Fähigkeit zur Bildung roter Blütenfarbe" geerbt, und das äußert sich darin, daß er eine wesentlich blassere Farbe aufweist.
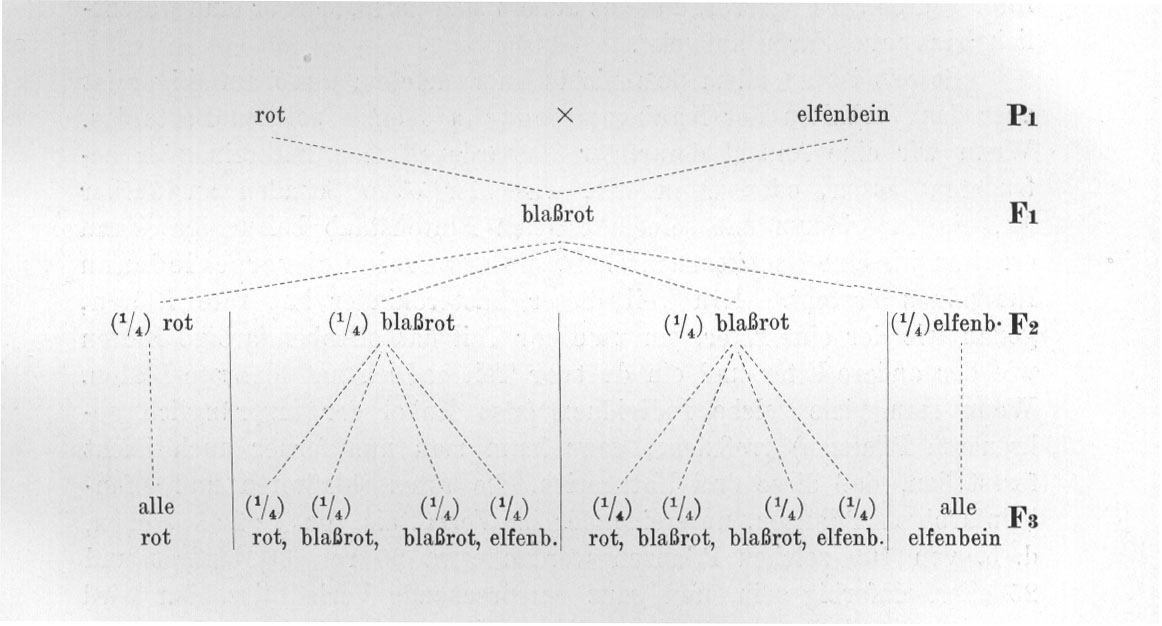
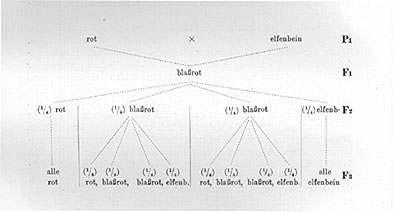
Soweit ist an allem dem nichts Unerwartetes; umso auffälliger ist aber das Verhalten der Nachkommenschaft eines solchen Bastardes. Wenn wir eine Anzahl derartiger Bastarde Ff sich untereinander befruchten lassen, oder wenn wir, was hier ohne Schaden ausführbar ist, ein Individuum mit seinem eigenen Blütenstaub befruchten, dann erhalten wir eine Nachkommenschaft, die aus dreierlei verschiedenen Individuen besteht. Ein Teil dieser Bastardkinder hat rote Blüten, genau wie der eine Elter, ein zweiter Teil hat elfenbeinfarbige Blüten wie der andere Elter und ein dritter Teil endlich hat blaßrote Blüten. Wenn man viele solche Individuen (also Enkel der ursprünglich gekreuzten Pflanzen) großzieht, dann kann man nun ferner auch leicht feststellen, daß diese drei Kategorien, die roten, blaßroten und elfenbeinfarbigen Pflanzen, untereinander im Verhältnis von 1 : 2 : 1 stehen, d. h. von 100 solchen Pflanzen werden etwa 25 rot, 50 blaßrot und 25 elfenbeinfarbig sein, und ganz entsprechende Verhältniszahlen wird man immer erhalten, wenn man diesen Versuch wiederholt. In einem konkreten Falle erhielt ich z. B. von 97 Pflanzen 22 rot, 52 blaßrot und 23 elfenbeinfarbig.
Die auf diese Weise gewonnenen roten Individuen erweisen sich in ihrer Deszendenz als völlig konstant, geben ausschließlich eine rote Nachkommenschaft, die elfenbeinfarbigen sind ebenfalls konstant, aber die blaßroten Individuen verhalten sich in ihrer Deszendenz genau wie der erste ursprüngliche Bastard Ff, d. h. "spalten" wieder auf in 1/4 rote, 2/4 blaßrote und 1/4 elfenbeinfarbige Nachkommen.
In Form eines Stammbaumes ist der Versuch auf Seite 48 dargestellt.
Man bezeichnet mit den hier an der rechten Seite beigedruckten Buchstaben Pl, Fl, F2 folgendes: Pl ist 1. Parentalgeneration, d. h. die ursprünglich zur Kreuzung verwendeten Individuen, F1 ist die 1. Filialgeneration, d. h. die primären Bastarde und F2 usw. sind die weiteren Generationen. Ganz entsprechend ist P2 die Elterngeneration von P1 usw.
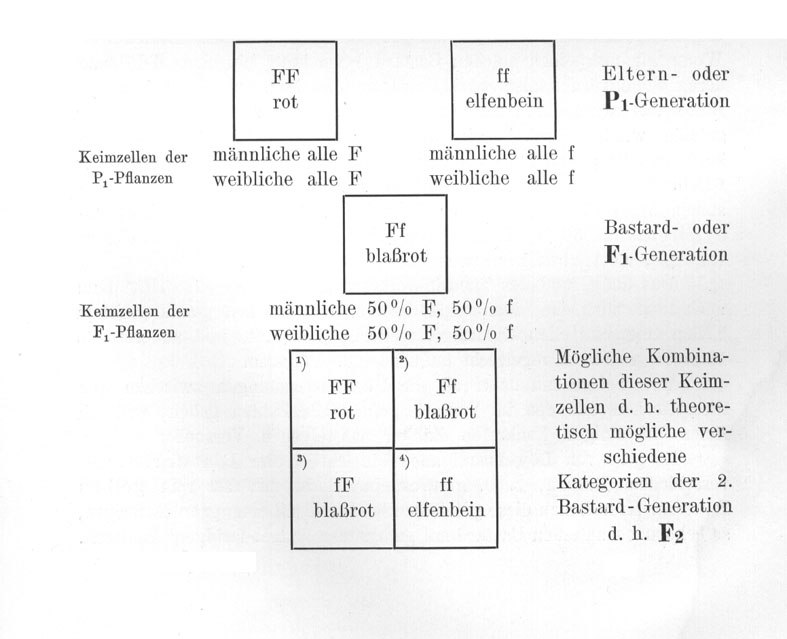
Wie kommt nun dies alles zustande? - Die Lösung gibt die zuerst von MENDEL aufgestellte, heute fast zur Gewißheit gewordene Hypothese, daß jeder derartige Bastard zweierlei Arten von Sexualzellen bilde und zwar 50% "väterliche" und 50% "mütterliche".
Die Konsequenz dieser Theorie wollen wir jetzt im einzelnen an der Hand unseres Beispieles besprechen. Der Antirrhinum-Bastard Ff auf Tafel II, Fig. 3 bildet nach der Theorie zweierlei Arten von Sexual-
zellen und zwar ist die eine Hälfte davon ganz genau von der Art F, die also genau ebenso wie die Sexualzellen einer homozygotischen [homozygoten] Pflanze FF die Fähigkeit zur Bildung roter Farbe übertragen, und die andere Hälfte seiner Sexualzellen ist genau gleich den Sexualzellen f der elfenbeinfarbigen Rasse ff. Diese Annahme, daß ein solcher Bastard zweierlei Sexualzellen bildet - zweierlei männliche sowohl wie zweierlei weibliche - und zwar zu je 50 %, ist der Kernpunkt der MENDELschen Theorie. Alles andere ist erst sekundär und unwichtig.
Wenn wir nun unsern Antirrhinum-Bastard Ff mit seinem eigenen Blütenstaub befruchten, oder wenn wir mehrere solche Bastarde einander gegenseitig befruchten lassen, dann können die beiden verschiedenen Kategorien F und f von Sexualzellen sich in vier verschiedenen Weisen kombinieren.
|
Eine Eizelle F kann treffen ein Pollenkorn F und gibt ein Individuum FF, das rot blüht, |
|
eine Eizelle F kann treffen ein Pollenkorn f und gibt ein Individuum Ff, das blaßrot blüht, |
|
eine Eizelle f kann treffen ein Pollenkorn F und gibt ein Individuum fF, das blaßrot blüht, |
|
eine Eizelle f kann treffen ein Pollenkorn f und gibt ein Individuum ff, das elfenbeinfarbig blüht. |
Da alle vier möglichen Kombinationen die gleiche Wahrscheinlichkeit haben, so werden wir erwarten dürfen, daß in der Deszendenz eines solchen Bastardes diese vier verschiedenen Arten von Individuen FF, Ff, fF und ff in annähernd gleichen Verhältnissen vorkommen werden, d. h. daß die Nachkommenschaft zusammengesetzt sein wird aus:
|
1/4 Individuen, welche entstanden sind als FF |
d. h. homozygotisch [homozygot] konstant rot sind, |
|
1/4 Individuen, welche entstanden sind als Ff
1/4 Individuen, welche entstanden sind als fF |
d. h. heterozygotisch [heterozygot] blaßrote Bastarde sind, |
|
1/4 Individuen, welche entstanden sind als ff |
d. h. homozygotisch [homozygot] konstant elfenbeinfarbig sind. |
In Form eines Schemas pflegt man einen derartigen Kreuzungsversuch folgendermaßen darzustellen:
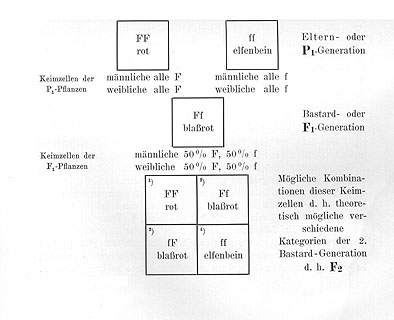
Wir sehen also, daß diese Zusammensetzung von [einer] F2 einer derartigen Kreuzung aus 1/4 rein mütterlichen, 1/4 rein väterlichen Individuen und 2/4 Bastarden auf Grund der Theorie erwartet werden muß. Die Theorie erklärt in sehr einfacher Weise die eigentümlichen Spaltungserscheinungen. Sie steht aber auch sonst so sehr im Einklang mit allen weiterhin gemachten Beobachtungen, daß wir mit ihr als etwas ganz sicher Begründetem arbeiten können.
Es ist möglich gewesen, auf Grund der Theorie nun auch das Resultat von weiteren Versuchen vorherzusagen und so die Theorie zu prüfen.
Einen Versuch, der seine Hypothese prüft, hat schon MENDEL selbst ausgeführt, nämlich die Rückkreuzung eines Bastardes mit dem einen seiner Eltern.
Wir wollen einen analogen Rückkreuzungsversuch mit unserem Löwenmaulbastard ausführen. Wir gehen in der Weise vor, daß wir unseren Bastard, der die Formel Ff hat, befruchten mit Blütenstaub der elfenbeinfarbigen Elternpflanze ff oder einer anderen elfenbeinfarbigen Pflanze der gleichen Sippe. Was ist da für ein Resultat zu erwarten, wenn wir uns auf den Boden der Spaltungshypothese stellen?
Nach der Hypothese produziert der Bastard zweierlei Eizellen, die eine Hälfte der Eizellen überträgt nur das Merkmal rote Blütenfarbe, die andere Hälfte überträgt nur das Merkmal elfenbein Blütenfarbe. Wenn wir nun einen solchen Bastard befruchten mit einer ff-Pflanze, deren Pollenkörner also sämtlich nur das Merkmal elfenbeinfarbige Blüte übertragen, dann müssen 50 % der so entstehenden Nachkommen gebildet werden durch Vereinigung einer F-Eizelle mit einem f-Pollenkorn und die anderen 50 % durch Vereinigung einer f-Eizelle mit einem f-Pollenkorn. Es müssen also bei einer solchen Rückkreuzung entstehen 50 % Pflanzen von der Formel Ff, d. h. blaßrote Heterozygoten und 50 % Pflanzen von der Formel ff, d. h. elfenbeinfarbige, weiterhin konstant bleibende Pflanzen.
Das auf Grund der Theorie vorherzusagende Ergebnis trifft denn auch tatsächlich ein. Man erhält im Versuche zu fast genau gleichen Teilen einerseits elfenbeinfarbige, weiterhin konstante und andererseits blaßrote, weiterhin regelrecht aufmendelnde Pflanzen.
Wie weitgehend dabei oft die Übereinstimmungen zwischen den theoretischen und den im Versuch gefundenen Zahlen gehen, will ich Ihnen an einigen konkreten Zahlen aus eigenen Versuchen zeigen. Es gibt von Löwenmaul auch hinsichtlich der Blattfarbe sehr verschiedene Rassen, z. B. gibt es eine Rasse mit fast rein gelben Blättern. Kreuzt man diese gelbblätterige Rasse mit einer grünblätterigen, so bekommt man einen Bastard mit gelbgrünen "aurea-farbigen" Blättern. Bei Selbstbefruchtung spaltet er auf in 1/4 grüne, 2/4 aureafarbige und 1/4 gelbe Keimlinge.
Wenn man nun solch einen Bastard mit dem grünen Elter rückkreuzt, dann ist auf Grund der Theorie zu erwarten, daß 50 % der so erhaltenen Pflanzen grün und weiterhin konstant und 50 % aureafarbig und weiterhin spaltend sein müssen. In einer entsprechenden Versuchsreihe bekam ich aus solcher Rückkreuzung zwischen den Bastarden und den grünen Eltern insgesamt 1178 Pflanzen, von denen 597 gelbgrün und 581 grün waren, also fast genau das theoretisch zu erwartende Verhältnis 1 : 1.
In unseren bisherigen Beispielen war es immer ohne weiteres leicht möglich, die Bastarde als solche an ihrer Farbe schon zu erkennen. Die Bastarde zwischen dem elfenbeinfarbigen und dem roten Löwenmaul, die Ff-Individuen, sind meist deutlich durch ein blasses Rot von den homozygotischen [homozygoten] Eltern verschieden, so ist es auf Taf. II leicht möglich, die FF- und Ff-Individuen zu unterscheiden. Die Bastarde nehmen hier also gewissermaßen eine Art Mittelstellung zwischen den Eltern ein. Das gleiche gilt auch für die Bastarde zwischen den beiden eben erwähnten, in der Blattfarbe verschiedenen Löwenmaulrassen. So ist die Sachlage aber durchaus nicht immer.
Das Aussehen der Heterozygoten kann auch ein ganz anderes sein. Sehr eigenartige Verhältnisse finden sich z. B. bei der Krenzung von zwei Hühnerrassen, über die BATESON (14, I-IV) berichtet hat. Andalusier-Hühner gibt es in verschiedenen konstanten Farbenrassen, u. a. kennt man eine Rasse, die schwarz und eine Rasse, die weiß mit schwarzen Flecken ist. Kreuzungen zwischen schwarzen Hühnern und schwarzweißen Hähnen und ebenso zwischen schwarzweißen Hühnern und schwarzen Hähnen geben nun immer Individuen, die blau gefiedert sind. Also hier zeigen die Heterozygoten eine ganz neue Farbe. Aber wenn man derartige blaue Andalusier aus F1 unter sich kreuzt, dann erhält man ganz regelmäßig eine F2, die aus 1/4 schwarzen, 1/4 schwarzweißen und 2/4 blauen Hühnern sich zusammensetzt. Das ist genau das, was auf Grund der Spaltungsgesetze erwartet werden mnß. Die schwarzen und die schwarzweißen Hühner erweisen sich bei Inzucht als konstant, die blauen spalten weiter in dem alten Verhältnis auf.
Allzu häufig scheint diese Sachlage, daß die Heterozygoten eine total von derjenigen der Homozygoten verschiedene Farbe aufweisen, nicht vorzukommen. Immerhin sind aber doch schon mehrere derartige Fälle genau bekannt. [Das sind die Ausnahmen geblieben.]
Ein dritter Fall, der ganz besonders häufig vorkommt, ist der, daß die Heterozygoten ganz dem einen Elter gleichen, z. B. wenn wir eine rote Löwenmaulpflanze kreuzen mit einer weißen (Fig. 1 Taf. 1), erhalten wir Bastarde, die nicht von den homozygotischen roten Individuen zu unterscheiden sind und dementsprechend besteht also äußerlich [die] F2 einer solchen Kreuzung aus 3/4 roten und 1/4 weißen Individuen. Aber eine Prüfung der Deszendenz dieser roten Individuen zeigt, daß auch hier ein Teil konstante Deszendenz hat, sich als homozygotisch [homozygot] erweist; zwei andere Teile dagegen auch weiterhin spalten. Also auch hier sind in Wirklichkeit die Verhältnisse die gleichen wie bei den Bastarden zwischen rot und elfenbeinfarbig.
Diese Erscheinung, daß vielfach die Bastarde äußerlich nicht von dem einen Elter zu unterscheiden sind, bezeichnet man mit dem Wort Dominanz. Man sagt, das eine Merkmal, hier etwa die Fähigkeit zur Bildung roter Farbe, "dominiere" über das andere Merkmal, über das Fehlen dieser Fähigkeit, und dieses letztere Merkmal sei "rezessiv" gegen das erstere. Man hat dieser Dominanzerscheinung, die durchaus keine allgemeine Regel ist, vielfach übertrieben große Bedeutung zugeschrieben, von einer "Dominanzregel" gesprochen. Das ist ganz verkehrt. Eine irgendwie allgemein gültige Dominanzregel gibt es nicht und sehr häufig ist eine Dominanz nur scheinbar. Es kann, darauf hat zuerst CORRENS (72) hingewiesen, für unser Auge ein Bastard völlige Dominanz, etwa in der Blütenfarbe eines Elters zeigen, aber wenn wir die Farbe des Bastards kolorimetrisch untersuchen, dann sehen wir, daß er, im Grunde genommen, sich so verhält, wie der Bastard zwischen dem roten, und dem elfenbeinfarbigen Antirrhinum, d. h. daß die Färbung des Heterozygoten viel schwächer ist, als die der Homozygoten, daß also die völlige Dominanz nur eine scheinbare ist.
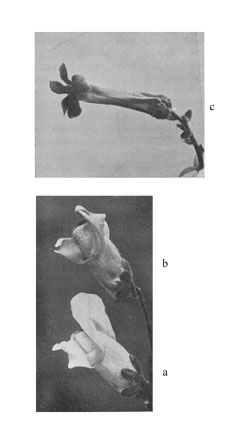
Fig. 14. Löwenmaulrassen (Antirrhinum). a Rasse mit normalen (zygomorphen) Blüten; c Rasse mit pelorischen (radiären) Blüten; b F1-Bastard zwischen a und c. Nahezu völlige Dominanz.
Ob überhaupt völlige Dominauz, d. h. die Erscheinung, daß ein Merkmal sich heterozygotisch [heterozygot] schon genau so äußert wie homozygotisch [homozyygot], häufig vorkommt, ist mir fraglich. Scheinbare völlige Dominanz beruht eben wohl oft nur auf unserem mangelhaften Unterscheidungsvermögen.
Die empirischen, in den Versuchen gefundenen Zahlen stimmen mit den theoretisch erwarteten auch in diesen Fällen von völliger oder nahezu völliger Dominanz genügend überein, wenn man nur mit der nötigen Individuenzahl arbeitet. Z. B. fand MENDEL selbst in seinen klassischen Kreuzungsversuchen mit Erbsenrassen folgende Zahlen: Er kreuzte eine Erbsenrasse mit grünen Keimblättern mit einer anderen mit gelblichen Keimblättern. Die Bastarde hatten alle gelbliche Keim-

Fig. 15. Löwenmaulrassen (Antirrhinum). a völlig normale Rasse; c Schlitzer-Rasse; b Bastard zwischen beiden, unterscheidet sich von a dadurch, daß die Zipfel der Unterlippe in der Mitte eingekerbt sind.
blätter, d. h. das Merkmal gelblich dominiert über das Merkmal grün. Von 258 solcher F1-Bastarde erhielt MENDEL 8023 Individuen der F2-Generation, und von diesen hatten 6022 gelbliche und 2001 grüne Keimblätter, d. h. die gelben stehen zu den grünen fast genau in dem theoretisch zu erwartenden Verhältnisse 3 : 1.
Hat man durch langes Arbeiten mit einem bestimmten Versuchstier oder einer Versuchspflanze seinen Blick geschärft, dann wird man wohl meistens Unterschiede zwischen den Homozygoten und den Heterozygoten erkennen. Fig. 14 und 15 zeigen das Aussehen zweier mendelnden Bastarde verglichen mit ihren Eltern. Man sieht, daß in Fig. 14 der Bastard (b) zwischen einer zygomorphen (a) und einer radiären (c) Rasse des Löwenmauls deutlich eine kleinere Oberlippe hat als die homozygotisch [homozygot] zygomorphe Elternrasse. In Fig. 15 erkennt man den Bastard (b) zwischen der normalen zygomorphen Rasse (a) und einer anderen mit eigentümlich mißbildeten Blüten (c) daran, daß die Zipfel der Blumenblätter des Bastardes alle noch ein weiteres Mal geteilt sind, sie zeigen alle in ihrer Mitte eine leichte Einkerbung.
Wenn nun auch freilich meistens die Heterozygoten äußerlich verschieden sind von den Homozygoten, so ist dennoch sehr häufig eine reinliche Unterscheidung, etwa bei Auszählen einer F2-Generation, nicht möglich. Es rührt daher, daß sehr oft die Homo- und die Heterozygoten transgredierend modifizierbar sind (vergl. Seite 31). Schon in unserem ersten Beispiel der Kreuzung eines elfenbeinfarbigen Löwenmauls mit einem roten hat man oft Schwierigkeiten, in [der] F2 die Homozygoten und Heterozygoten zu unterscheiden. Schattig stehende Homozygoten haben ebenfalls ein etwas blasses Rot und sind nicht immer mit Sicherheit von Heterozygoten zu unterscheiden, die unter Verhältnissen aufgewachsen waren, welche eine dunklere Rotfärbung verursachen.
Die Dominanzverhältnisse sind auch bei einem und demselben Individumn öfters ungleich, es kann in der Jugend ein Tier deutliche Dominanz eines einelterlichen Merkmales zeigen, während in alten Stadien diese Dominanz nicht mehr zu erkennen ist. Z. B. hat A. LANG bei Schnecken-Kreuzungen ähnliches beobachtet.
___________________
Kreuzungsversuche wie die bisher genannten sind in den letzten Jahren in sehr großer Zahl mit den verschiedensten Organismen angestellt worden. Man hat gefunden, daß überall die gleichen Gesetzmäßigkeiten in Geltung sind, einerlei, ob wir mit Löwenmäulchen oder mit Getreidearten, mit Mäusen oder Käfern, mit Schnecken oder Schmetterlingen experimentierten. Wir finden überall, daß, wenn die gekrenzten Rassen sich in einem Merkmal unterscheiden, dann die Bastarde immer so aufspalten wie der Löwenmaulbastard unseres Schulbeispiels, d. h. daß also [die] F2 zusammengesetzt ist aus 2/4 Individuen, die wieder Bastarde sind, und je 1/4 Individuen, die genau das väterliche bzw. mütterliche Merkmal aufweisen.
___________________
Genau die gleichen Gesetze gelten nun auch mutatis mutandis, wenn wir zwei Rassen kreuzen, die sich in mehr als einem Merkmal unterscheiden. Wir wollen uns hier auch zur Orientierung einen möglichst einfachen und übersichtlichen Fall aussuchen, ebenfalls wieder eine Löwenmaulkreuzung. Die Löwenmaulrassen unterscheiden sich z. B. auch in Merkmalen der Blütenform. So besitze ich eine völlig konstante Rasse, die nur Blüten trägt wie die in Fig. 14c abgebildete. Die Blüten sind hier völlig radiär gebaut, alle Blumenblätter haben die

Form, die in der normalen, zygomorphen Löwenmaulblüte allein das mittlere Blumenblatt der Unterlippe aufweist, und sind auch alle wie dieses kurz gespornt. Diese pelorische Rasse ist völlig fruchtbar und kann mit normalen Rassen gekreuzt werden. Tafel III stellt die Kreuzung dieser roten pelorischen mit einer konstanten normal blühenden elfenbeinfarbigen Rasse dar. Der Bastard, die Fl-Pflanze, ist blaßrot

Fig. 16. Hochwüchsige und niedrigwüchsige Rasse vom Gartenlöwenmaul (Antirrhinum). Gleich alte und gleich behandelte Pflanzen.
und hat völlig normale Blütenform, nur die Oberlippe ist etwas kleiner als bei den homozygotisch [homozygot] normalen Sippen. Wir haben also hier einen Fall fast völliger Dominanz der normalen Form über die pelorische, während hinsichtlich der Farbe der Bastard eine ungefähre Mittelstellung einnimmt, wie das ja auch in den früher besprochenen Beispielen einer Kreuzung von rot mit elfenbeinfarbig der Fall war.
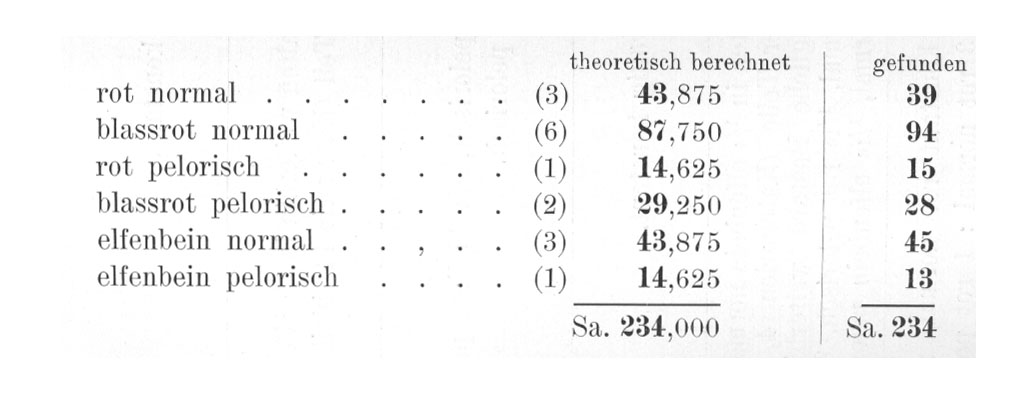
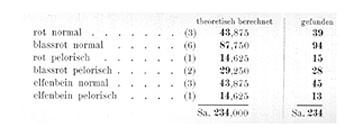
Die durch Selbstbefruchtung eines solchen Bastardes gewonnene F2-Generation zeigt, wie Tafel III veranschaulicht, eine sehr bunte Zusammensetzung. Es treten sechs äußerlich verschiedene Kategorien von Pflanzen auf, nämlich folgende:
|
Rote normale |
39 |
|
blaßrote normale |
94 |
|
rote pelorische |
15 |
|
blaßrote perlorische |
28 |
|
elfenbeinfarbige normale |
45 |
|
elfenbeinfarbige pelorische |
13. |
Das Zahlenverhältnis, in dem in einem meiner Versuche (S. 56, 1909) diese verschiedenen Kategorien gefunden wurden, ist in der Tabelle beigefügt.
Wie auf Grund der Spaltungsgesetze diese Kategorien und diese Verhältniszahlen zu verstehen sind, wollen wir in ähnlicher Weise ableiten, wie wir es vorhin für die Kreuzungsversuche mit den beiden nur in der Blütenfarbe verschiedenen Löwenmaulrassen getan haben.
Eine Sexualzelle oder wie man jetzt in der Vererbungsliteratur meistens sich ausdrückt, einen "Gameten" der normalen elfenbeinfarbigen Rasse wollen wir bezeichnen mit fE, einen Gameten der roten pelorischen Rasse mit Fe. Dabei soll bedeuten:
|
F |
Fähigkeit zur Bildung neuer Blütenfarbe. |
|
f |
Fehlen dieser Fähigkeit, d. h. der so bezeichnete Gamet überträgt elfenbeinfarbige Blüte. |
|
E |
Fähigkeit zur Bildung normaler Blüten. |
|
e |
Fehlen dieser Fähigkeit, d. h. der so bezeichnete Gamet überträgt pelorische Blütenform. |
Die aus der Vereinigung zweier Gameten Fe (rote Blütenfarbe, pelorische Blüten) entstandene P1-Pflanze als der roten pelorischen Rasse bekommt also die Formel FFee.
Die aus der Vereinigung zweier Gameten fE (elfenbeinfarbige, normale Blüte) entstandene P1-Pflanze aus der elfenbeinfarbigen normalen Rasse bekommt die Formel ffEE.
Der Bastard hat dann natürlich die Formel FfEe, da er ja durch die Vereinigung eines Gameten Fe und eines Gameten fE entstanden ist.
Wenn nun dieser Bastard geschlechtsreif wird, dann bildet er nach der MENDELschen Spaltungshypothese im ganzen viererlei verschiedene Gameten aus, nämlich folgende:
|
FE |
Die rote Farbe und normale Form übertragen. |
|
Fe |
Die rote Farbe und pelorische Form übertragen. |
|
fE |
Die elfenbein-Farbe und normale Form übertragen. |
|
fe |
Die elfenbein-Farbe und pelorische Form übertragen. |
Diese vielerlei verschiedenen Gameten werden alle in ungefähr gleicher Zahl gebildet, also jede dieser vier Kategorien bildet etwa 1/4 der sämtlichen Gameten, der männlichen sowohl der weiblichen.
Wenn wir nun einen solchen Bastard FfEe mit sich selbst, oder mehrere solcher Bastarde gegenseitig miteinander befruchten, dann können sich diese vier Kategorien von weiblichen Sexualzellen mit den vier Kategorien von männlichen Sexualzellen in 16 verschiedenen Kombinationen vereinigen, nämlich:

In Form eines vielfach zur Illustrierung derartiger Versuche gebrauchten Schemas stellt sich diese Spaltung und Neukombination der Merkmale folgendermaßen dar.
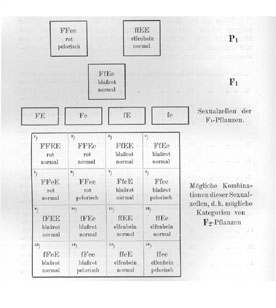
Ein Blick auf diese Übersicht zeigt, daß von den 16 möglichen Kombinationen
drei (1, 2, 5) Pflanzen mit roten normalen,
sechs (3, 4, 7, 9, 10, 13) Pflanzen mit blaßroten, normalen,
eine (6) Pflanze mit roten pelorischen,
zwei (8, 14) Pflanzen mit blaßroten pelorischen,
drei (11, 12, 15) Pflanzen mit elfenbeinfarbigen normalen,
eine (16) Pflanzen mit elfenbeinfarbigen pelorischen
Blüten ergeben müssen.
Wenn wir die blaßroten und die roten Individuen nicht unterscheiden, sondern zusammenzählen, dann werden wir finden, daß
|
9 |
Kategorien rot normal (beide Merkmale in der dominierenden Form), |
|
3 |
Kategorien rot pelorisch (nur ein Merkmal in der dominierenden Form), |
|
3 |
Kategorien elfenbeinfarbig, normal (nur ein Merkmal in der dominierenden Form), |
|
1 |
Kategorien elfenbeinfarbig pelorisch (kein Merkmal in der dominierenden Form), |
sein werden.
Da alle Kombinationen die gleiche Wahrscheinlichkeit haben, ist auf Grund der Theorie zu erwarten, daß von F2-Pflanzen einer solchen Kreuzung:
|
3/16 |
rot normal blühen müssen, |
|
6 /16 |
blaßrot normal blühen müssen, |
|
1 /16 |
rot pelorisch blühen müssen, |
|
2 /16 |
blaßrot pelorisch blühen müssen, |
|
3 /16 |
elfenbein normal blühen müssen, |
|
1 /16 |
elfenbein pelorisch blühen müssen, |
Vergleichen wir damit die im Versuche gefundenen Zahlen, so finden wir eine sehr weitgehende Übereinstimmung:
Auf Grund der Theorie ist nun ferner zu erwarten, daß je 1/3 der normal blühenden Kategorien weiterhin konstant normal blühende Deszendenz haben wird, nämlich alle Individuen aus EE-Kategorien (Nr. 1, 3, 9, 11), ferner werden wir erwarten müssen, daß je 2/3 aller Individuen, nämlich aus den Kategorien Ee (Nr. 2, 4, 5, 7, 10, 12, 13, 15) weiterhin aufmendeln müssen in 1/4 normale und 3/4 pelorische Pflanzen. Auch dieses theoretische Postulat wird duch den Versuch erfüllt. Tatsächlich erweist sich in allen derartigen Kreuzungen ein entsprechender Teil der normalen F2-Pflanzen, der übrigens auch an der kleineren Oberlippe meist äußerlich kenntlich ist, als Ee, d. h. mendelt in normale und pelorische Individuen auf.
Wenn wir auch diesen Versuch in Form eines Stammbaumes darstellen, bekommen wir:

Wir sehen also, daß die verschiedenen Merkmale, durch welche die beiden ursprünglich gekreuzten Rassen sich unterscheiden, ganz unabhängig voneinander sich auf die Gameten des Bastardes verteilen, oder, wie man zu sagen pflegt, ganz unabhängig voneinander "mendeln". Dieses Gesetz der Selbstständigkeit der Merkmale ist neben der Spaltungsregel die wichtigste Grundlage der experimentellen Vererbungslehre. Auch dieses Gesetz hat schon MENDEL gefunden und mit aller Schärfe präzisiert.
Diese Erkenntnis hat eine große praktische Bedeutung besonders für die Pflanzen- und Tierzüchtung. Nehmen wir einmal an, pelorische Blüten und elfenbeinfarbige Blüten seien wünschenswerte Eigenschaften, eine solche Rasse existiere aber nicht, sondern nur zwei andere Rassen, die elfenbeinfarbige normale und die rote pelorische, die also jeweils nur die eine der beiden gewünschten Eigenschaften aufweisen. Ein Blick auf die Tafel III zeigt, daß man nur die beiden Rassen zu kreuzen braucht, um in der übernächsten, der F2-Generation, dann Individuen zu erhalten, welche diese beiden Eigenschaften in sich vereinigen und konstant auf ihre ganze Deszendenz vererben.
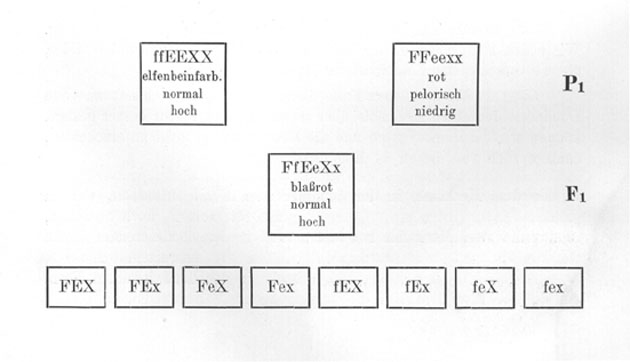
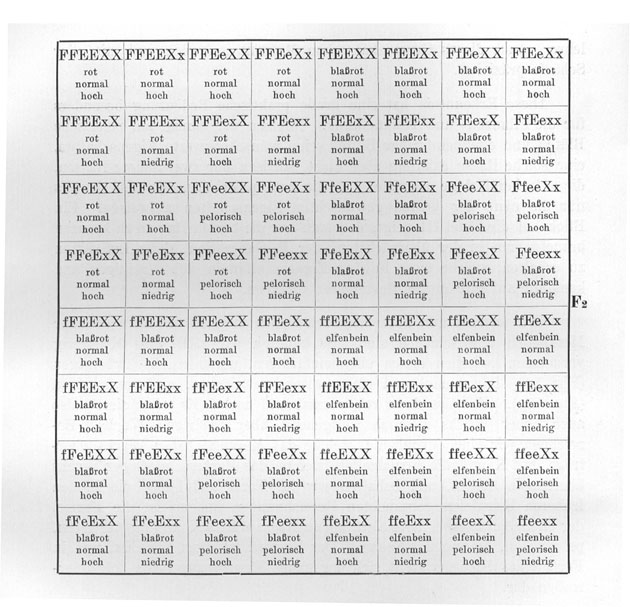
Wir können nun natürlich auch Sippen kreuzen, die in noch mehr Merkmalen unterschieden sind. So könnten wir z. B. eine Kreuzung vornehmen zwischen einem elfenbeinfarbigen, normalen, hochwüchsigen (Fig. 16b) Löwenmaul und einem roten, pelorischen, niedrigwüchsigen (Fig. 16a). [Die] Fl wäre dann rot, normal und hoch. Es "dominiert" also auch hoher Wuchs ziemlich weitgehend über niedrigen Wuchs. Bezeichnen wir eine Sexualzelle, welche das Merkmal hoher Wuchs überträgt, mit X und die Sexualzellen, welche das Merkmal niedriger Wuchs übertragen, mit x, und behalten wir für Form und Farbe die in den früheren Beispielen gebrauchten Bezeichnungen bei, so ist die Formel der elfenbeinfarbigen, normalen, hohen Rasse ffEEXX und die der roten, pelorischen, niedrigen FFeexx. Der Bastard ist dann FfEeXx, und in Form eines Schemas stellt sich der ganze Kreuzungsversuch folgendermaßen dar .
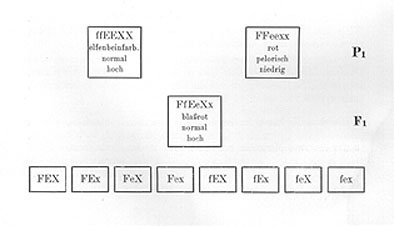
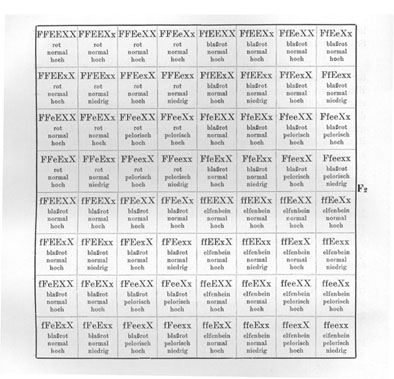
Eine ausführliche Legende zu geben ist wohl nicht mehr notwendig. Wir sehen also, daß hier der Bastard 8 verschiedene Sexualzellen bildet, die 64 verschiedenen Kombinationen in F2 ermöglichen.
Liegt in einem solchen Falle überall volle oder doch nahezu volle Dominanz des einen Merkmals über das andere vor, oder besser gesagt, können wir die Homozygoten und die Heterozygoten nicht unterscheiden, dann werden von diesen 64 Kombinationen
|
27 |
alle drei Merkmale in der dominierenden Form aufweisen, werden also rot, normal, hoch aussehen, |
|
9 |
nur die zwei Merkmale rot und normal in der dominierenden Form aufweisen, |
|
9 |
nur die zwei Merkmale rot und hoch in der dominierenden Form aufweisen, |
|
9 |
nur die zwei Merkmale normal und hoch in der dominierenden Form aufweisen, |
|
3 |
nur das eine Merkmale rot in der dominierenden Form aufweisen, |
|
3 |
nur das eine Merkmale normal in der dominierenden Form aufweisen, |
|
3 |
nur das eine Merkmale hoch in der dominierenden Form aufweisen, |
|
1 |
kein dominierendes Merkmal zeigen. |
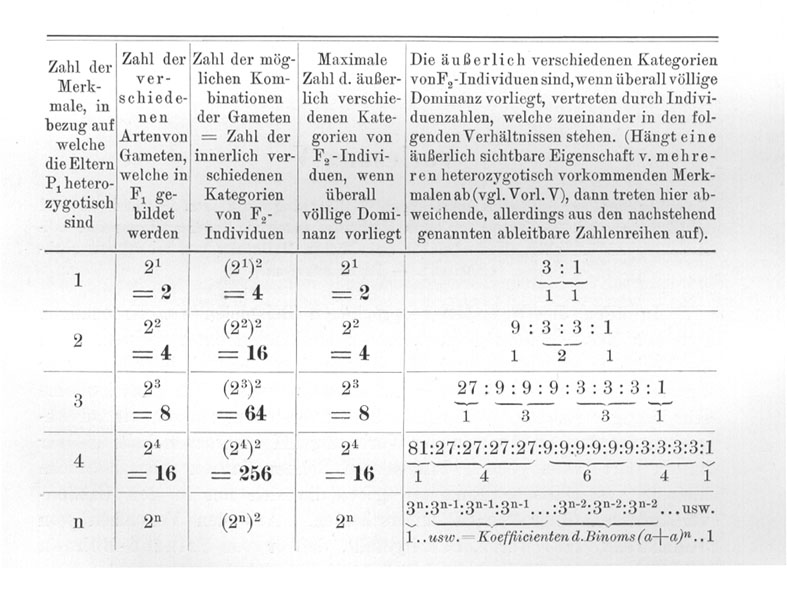
Kreuzen wir Sippen, die sich in noch mehr Merkmalen unterscheiden, dann gelten diese selben Gesetzmäßigkeiten weiter. Im einzelnen darauf einzugehen, ist wohl überflüssig. Was für Zahlenverhältnisse auftreten, und welche allgemeine Formel für n-Merkmale leicht aufgestellt werden kann, zeigt die nachstehende Tabelle ohne weitere Erklärung.
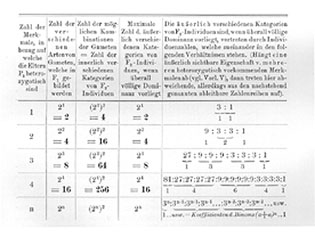
Vielleicht ist es Ihnen aufgefallen, daß der größte Teil aller bisher besprochenen Beispiele von Vererbungen nach den Spaltungsgesetzen sich auf Färbungsmerkmale bezog. Das rührt daher, daß Färbungsunterschiede im allgemeinen in Vererbungsversuchen viel leichter verfolgt werden können als andere Unterschiede, und liegt nicht etwa daran, daß nur gerade für die Färbungsvererbung die Spaltungsgesetzte gelten. Man hat bei Vererbungsversuchen natürlich mit Vorliebe auf Merkmale geachtet, die leicht erkennbar und augenfällig sind. Es ist sehr leicht, in einem Kreuzungsversuch zwischen einem roten und einem weißen Löwenmäulchen in F2 die Spaltung zu verfolgen, aber es ist sehr umständlich, die Spaltung festzustellen, wenn man z. B. kreuzt eine selbstfertile mit einer selbststerilen Rasse, wo man den F2-Pflanzen ja nicht ansehen kann, ob sie selbstfertil oder selbststeril sind, sondern wo man das erst wieder für jede einzelne F2-Pflanze durch einen eigenen Versuch feststellen muß. Eine Auszählung einer F2-Generation macht daher eine heillose Arbeit, wenn man die Vererbung derartiger Merkmale untersucht.
Also nur der Bequemlichkeit halber ist soviel gerade mit Farbenrassen experimentiert worden. Die Spaltungsgesetze gelten aber auch für alle anderen Merkmale ebenso gut wie für die Färbung.
Internet address
of this
document:
internetlibrary.html
© 2002 by Wolf-Ekkehard Lönnig - loennig@mpiz-koeln.mpg.de
 Disclaimer
Disclaimer