IV. 3. DER GENETISCHE ARTBEGRIFF
STÄRKEN
Der genetische Artbegriff schien gegenüber dem morphologischen und darwinistischen Artbegriff den entscheidenden Vorteil zu haben, dass das erste Mal ein naturwissenschaftlich fassbares, klar abgrenzbares und jederzeit reproduzierbares Kriterium von höchster biologischer Wertigkeit zur Grundlage der Artdefinition gemacht wurde: Die Kreuzbarkeit mit fertiler Nachkommenschaft.
An dieser Stelle sei nochmals an den Grundsatz von de Vries erinnert, "dass Formen, welche bei gegenseitigen Kreuzungen in allen Merkmalen den MENDEL'schen Gesetzen folgen, als Varietäten einer selben Art aufzufassen sind" (vgl. p. 57 dieser Arbeit; vgl. auch die Zitate von Nilsson und Lamprecht pp. 44-46).
Es kann überhaupt keinen Zweifel geben, dass der genetische Artbegriff ein ganz entscheidender Schritt in die richtige Richtung zu einem naturwissenschaftlich-objektiven Artbegriff ist. Jetzt entscheiden nicht mehr die morphologischen Spezialisten oder (neo-)darwinistischen Theoretiker mit ihren häufig reversiblen 'Artbarrieren', ob ein Merkmal, Merkmalskomplex oder eine Verhaltensweise bereits als Artkriterium gelten kann oder nicht (wobei der eine Sachbearbeiter die Frage im konkreten Fall unbedingt bejaht, der nächste im selben Fall aber mit Nachdruck verneint und sich so der Streit über mehrere Forschergenerationen hinziehen kann), sondern die naturwissenschaftliche Frage wird mit naturwissenschaftlichen Mitteln entschieden: die Kreuzbarkeit und reziproke Überführbarkeit der Merkmale.
Alle Formen, die eine reale oder potentielle Fortpflanzungsgemeinschaft bilden, gehören zur selben Art und seien sie morphologisch-physiologisch und ethologisch auch noch so verschieden, wie wir das für die Menschen-, Tauben-, Erbsenrassen etc. aufgeführt haben. Der genetische Artbegriff ist weitgehend mit der ursprünglich von Mayr gegebenen Artdefinition identisch: "Species are groups of actually or potentially interbreeding natural populations which are reproductively isolated from other such groups" (vgl. p. 38). Wie wir oben aufgeführt haben, hat Mayr 1969 das "or potentially" aus der Definition gestrichen, aber schon 1963/1967 wehrt er sich entschieden gegen den genetischen Artbegriff, wenn er 1967, p. 323 schreibt:
Einige Botaniker anerkennen eine besondere Artenkategorie für alle Populationen und Arten, die sich erfolgreich kreuzen. CLAUSEN, KECK und HIESEY (1945) haben als Cänospezies neu definiert, "alle die Ökospezies, die so verwandt sind, daß sie untereinander in einem begrenzten Ausmaß Gene durch Bastardierung austauschen können". Wenn es auch nicht zu bezweifeln ist, daß mit der Festigung vollständiger Kreuzungssterilität der Punkt totaler Irreversibilität der Speziation erreicht ist, so muß doch nachdrücklich betont werden, daß Sterilität nur einer der vielen isolierenden Mechanismen ist. Die Anerkennung der Cänospezies übertreibt die Wichtigkeit der Sterilitätsschranke gröblich und würde in der Systematik der Tiere zu absurden Schlüssen führen. Es ist bei Tieren nicht ungewöhnlich, daß alle Mitglieder einer Gattung oder sogar Unterfamilien miteinander Gene austauschen können, obwohl das in der Natur selten oder niemals vorkommt. Eine so reich gegliederte Gruppe wie die Schwimmenten (Anatini) in eine einzige Cänospezies zusammenzufassen, wäre sinnlos. In anderen Tiergruppen (Drosophila z.B.) können beträchtliche Schranken für den Genaustausch schon auf der Stufe der Subspezies vorhanden sein, und die meisten realen Arten sind miteinander steril. Ich sehe nichts, was die Anerkennung einer Artkategorie für Tiere empfehlen könnte, die der Cänospezies von TURESSON und seiner Schule entspricht.
Die Polemik gegen die Cänospezies als "gröbliche" Übertreibung der Sterilitätsschranke mit "absurden Schlüssen" und "sinnlosen" Zusammenfassungen ist ungerechtfertigt. Warum sollen eigentlich nicht alle Schwimmenten zu ein und derselben Art gehören? Für jemanden, der die Mendel'schen Regeln verstanden und in experimentellen Studien erlebt hat, was alles an erstaunlich unterschiedlichen Merkmalen, Merkmalskomplexen und Verhaltensprogrammen mühelos rekombinierbar ist (vgl. z.B. Mendel 1866; Bateson 1909, 1913; Gray 1958, 1971; Feddersen 1978; Gottschalk 1978/1984; Lönnig 1982, 1985), für den bestehen nicht die geringsten Schwierigkeiten, die Vielfalt der Schwimmenten zu einer Mendelart zusammenzufassen (Scherer und Hilsberg haben 1982 eine ausführliche Studie über die Kreuzbarkeit der Arten und Gattungen der Anatidae vorgelegt, die diesen Zusammenhang weiter verdeutlicht.) Hier sei auch wieder an die Rekombinationsquadrate pp. 80, 122, 125 erinnert, sowie an die Tatsache, dass sich innerartliche Rekombination auf allen Ebenen - morphologisch, physiologisch und ethologisch - abspielt.
Welche Schwierigkeiten Neodarwinisten selbst mit der konsequenten Anwendung ihres eigenen, durch die oben diskutierten nicht-genetischen Barrieren eingeschränkten Artbegriff haben können, zeigt Mayrs Kommentar zu Hybrid-Populationen beim Thema "Zufalls- oder selektive Paarung" (1967, p. 299):
Hatten sich erste Isolationsmechanismen vor der Verschmelzung entwickelt, werden deutliche Abweichungen von der Zufallspaarung zu erwarten sein. Bei den Spechten der Gattung Sphyrapicus fand HOWELL (1952) in einem nordkalifornischen Bastardgürtel acht offensichtlich reine nuchalis, 14 reine dagetti und 20 Bastarde. Wo die Verbreitungsgebiete von nuchalis und ruber sich in Kersley, Britisch Columbien, treffen, wird die eine Form durch die andere innerhalb der kurzen Strecke von 1,5 Meilen ersetzt. Von den in diesem Gebiet beobachteten Paaren schienen fünf nuchalis, drei ruber und drei Paare entweder gemischt ruber x nuchalis oder nuchalis x Hybride oder beide Partner Bastarde zu sein. Für weitere Fälle siehe A. MILLAR (1956) und SIBLEY (1959). Wo die beiden Paradies-Elstern Astrapia stephaniae und A. mayeri sich in Neu-Guinea im Gebiet des Hagengebirges treffen, tritt beträchtliche Bastardierung auf, doch ist auch das Fortbestehen der Elterntypen belegt, vielleicht als Ergebnis von Paarungsbevorzugung (MAYR und GILLIARD 1952 a). In solchen Fällen eines nur teilweisen Zusammenbruchs der Fortpflanzungsisolation ist ein Entscheid über die taxonomische Bewertung (Art oder Unterart) sehr schwierig.
In den meisten Fällen sekundärer Intergradation hat die genetische Divergenz noch nicht den Punkt erreicht, wo Paarungsbevorzugung auftritt.
Man sieht an diesem Beispiel, wie sehr der Autor noch dem morphologischen Artbegriff verhaftet ist (vgl. auch p. 18). So stark differierende Formen, wie wir sie bei den Paradieselstern Neuguineas vorfinden, können ja nicht zu einer Art gehören, selbst wenn keinerlei wie auch immer geartete Barrieren nachweisbar und die unterschiedlichen Populationen bestimmter Rekombinanten durch Hybridpopulationen miteinander verbunden sind.
Genetisch betrachtet ist die Situation in solchen Fällen jedoch völlig klar: es handelt sich um eine einzige Mendelpopulation, und alle wie auch immer beschaffenen Rekombinanten einer tatsächlichen oder potentiellen Mendelpopulation gehören auch zur gleichen Art.
Werfen wir in diesem Sinne einen Blick auf die Galapagos-Finken. Derzeit werden bis zu 6 Gattungen mit insgesamt 14 Arten beschrieben. Stellt sich jedoch die Art "als Summe ähnlicher miteinander kreuzbarer Individuen dar" (Kämpfe und Günther 1980/1985, p. 56), so dürften sich die Gattungs- und Artenzahlen stark reduzieren: B.R. Grant und P.R. Grant z.B. berichten 1982, dass der Dickschnabel-Grundfink (Geospiza magnirostris) und der Große Kaktusfink (G. conirostris) sowie der Mittlere Grundfink (G. fortis) und der Kleine Grundfink (G. fuliginosa) auf den Galapagos-Inseln Genovesa bzw. Daphne hybridisieren und "viable offspring" hervorbringen (- ein Punkt, der schon von Lack 1940 aufgrund seiner morphologischen Geospezinae-Studien wahrscheinlich gemacht worden war; vgl. auch Mayr 1942).
[P.R. Grant (1986, p. 353): "Intersterility is not known in Darwin's finches. Intrageneric hybrids among ground finches are certainly both viable and fertile (chapter 8), and probably the same is true for intergeneric hybrids between tree finches and warbler finches..." (Ecology and evolution of Darwin's finches. Princeton.)
Neil weist (1987, p. 245; Molecular evolutionary genetics; New York) auf die geringe genetische Distanz innerhalb von Vogelfamilien hin: "...six species of Geospiza (finches) in the Galapagos Islands show a genetic distance ranging from 0.004 to 0.065." Bei den Menschenrassen liegen die Werte zwischen 0.01 und 0.03.]
Und Curio berichtet über seine Studien an den Darwinfinken u.a. 1980, p. 344:
Trotz zahlreicher Unterschiede sind diese im Laufe hinreichend langer Trennung auf den einzelnen Inseln hervorgegangenen Arten fortpflanzungsbiologisch doch nicht soweit voneinander entfernt; das beweisen die häufigen Mischungen zwischen Arten und Gattungen. Außerdem füttern zum Beispiel selbst die ausgeprägtesten Insektenesser unter den Darwinfinken ihre Jungen aus dem Kropf, wie es Körneresser tun, nicht aber aus dem Schnabel, wie es sonst bei den insektenessenden Familien stets der Fall ist.
Wenn es zwischen Arten und Gattungen 'häufige Mischungen' gibt, dann liegt die Annahme nahe, dass wir es auch hier genetisch nur mit einer Art zu tun haben (vgl. auch Goldschmidt, zitiert p. 102).
In diesem Zusammenhang sei an die große Variabilität der Schnabelgrößen und -formen unserer Haustauben erinnert (vgl. p. 28) - Differenzen, die bei vielen Formen in nur wenigen Jahrhunderten aufgetreten und zum Teil weitergezüchtet worden sind. Niemand hat aufgrund der Schnabelunterschiede und der z.T. recht erheblichen anatomischen und ethologischen Differenzen bei den Haustaubenrassen eigene Arten unterschieden. [T.B. Smith weist 1987 in seinem Artikel "Bill size polymorphism and intraspecific niche utilization in an African finch" (Nature 329, 717-719) auf einen weiteren Fall intraspezifischer starker Schnabelvariation hin: "Differences in bill size between morphs exceed those reported for several sympatric species of Darwin's finches believed to show character displacement, and like many co-occurring congeneric species the morphs differ in diet. ...Evidence that large- and small-billed forms can interbreed has also been reported." (P. 717)]
Kommen wir auf eine weitere schon mehrfach zitierte Tiergruppe zurück: Nach Hinweis, dass die drei wichtigsten Verwandten unserer Haushunde (Wölfe, Koyoten und Goldschakale) alle miteinander fruchtbare Nachkommen hervorbringen können, schreibt Trumler 1981, pp. 43/44:
Also müßten wir Wölfe, Koyoten und Goldschakale einer einzigen Art zurechnen - ein Vorschlag, den übrigens Hans von Lengerken vor vielen Jahren schon gemacht hat. Er ist aber nicht realisierbar, denn die drei Arten sind nun einmal in vielen Merkmalen unterschiedlich. In Europa und Asien fressen die Wölfe ihre nächsten Verwandten, die Schakale. In Nordamerika fressen die Wölfe ihre dortigen nächsten Verwandten, die Koyoten. Nur ganz im Süden, in Texas und Umgebung, da fressen die Wölfe die Koyoten nicht, sondern verpaaren sich mit ihnen, und diese Bastarde bekommen auch wieder Junge. So ist dort im Süden Nordamerikas, eine undurchschaubare Mischpopulation entstanden...*
*The red wolf vanished from the forests of America some 10 years ago, a victim of the destruction of its habitat and of crossbreeding with coyotes. (Scientific American 256, p. 48 (1987))
Und nach Aufführung aller möglichen Kreuzungen zwischen Goldschakal, Pudel, Dingo, Koyoten etc. stellt er noch fest (p. 44):
Frei streunende "Coy-dogs" haben einigen Zoologen und der auf Sensationen ausgehenden Presse der USA lange Zeit Rätsel aufgegeben, weil sie es - wie auch manche deutsche Forscher - nicht glauben wollten, daß sich Haushund und Koyote vermischen können.
Der Kreis rundet sich ab, wenn wir an die Wolf-Koyotenbastarde im Süden der Vereinigten Staaten denken. Er schließt sich ganz, wenn wir berücksichtigen, daß sich Dingos sowohl auf freier Wildbahn als auch in Gefangenschaft unbegrenzt mit Haushunden fortpflanzen.
Und trotzdem - kein Zoologe wird es wagen, Wolf, Koyote und Goldschakal als Unterarten einer Art zu bezeichnen. Es geht nicht.
Warum sollte das nicht gehen? 'Die drei Arten sind nun einmal in vielen Merkmalen unterschiedlich.' Warum geht das jedoch bei den rund 450 Rassen unseres Haushundes, die sich in vielen Fällen wesentlich stärker voneinander unterscheiden als die zitierten drei 'Arten'? Deutsche Schäferhunde z.B. haben schon wiederholt kleine Hunde wie Yorkshire Terrier getötet (ein Fall ist mir aus unmittelbarer Nachbarschaft bekannt). Dennoch reichen hier die Unterschiede im Körperbau, Fellbeschaffenheit und Verhalten nicht aus, eigene Arten zu unterscheiden.
Warum unterscheiden wir weniger differente Populationen in der Natur als eigene Arten, aber die stark unterschiedlichen Haustierformen nur als verschiedene Rassen, wenn doch nach Darwins Auffassung die Haustierzucht das große Zuchtexperiment in der Natur belegen soll? In diesem Falle müsste es doch gerade umgekehrt sein: die weniger differenten Formen müssten als Unterarten und stärker voneinander abweichende Formen des Hausstandes als volle Arten geführt werden.
Auf genetischer Ebene stellen sich solche Probleme nicht. Alle Formen sind miteinander kreuzbar, alle Formen können fruchtbare Nachkommen miteinander haben. Es handelt sich um eine Mendelpopulation und damit auch bei den unterschiedlichsten Angehörigen dieses Formenkreises ausnahmslos um die Vertreter derselben Art, ob in der freien Natur oder als Haustiere. Mit Artbildung hat das Ganze nichts zu tun.
Wenden wir uns der Botanik zu:
Ganders und Nagata berichten über ihre und Gillets Kreuzungsstudien der hawaiischen 'Arten' der Kompositengattung Bidens (1984, pp. 182, 183, 191):
Gillet (1975) obtained fertile hybrids among 11 of the 19 species and 2 of the subspecies of Hawaiian Bidens. We have extended this crossing program so that hybrids have now been obtained involving all Hawaiian Species and all subspecies except B. hillebrandiana ssp. hillebrandiana, B. micrantha ssp. kalealaha and B. campylotheca ssp. waihoiensis (Table I). All hybrids that have flowers so far are completely pollen and seed fertile.
All Hawaiian Bidens appear to be interfertile so that interfertility cannot be used as a criterion for species delimitation, unless one is willing to consider them all a single species which exhibits greater morphological diversity than the rest of the genus. ...We have used ...a species concept based on morphological characters and ecological considerations.
In summary, the 19 species and eight subspecies of Bidens endemic to the Hawaiian Islands evolved from a single ancestral species. Morphologically they are more diverse than the Bidens on any continent, but the Hawaiian species are interfertile.
Außerdem heben die Autoren hervor, dass Bidens cosmoides nach Gillets Angaben nicht mit anderen Linien gekreuzt werden konnte, sie selbst aber keine Schwierigkeiten mit dieser Frage hatten: "We found that B. cosmoides would cross freely with other species" (p. 182).
"...unless one is willing to consider them all a single species which exhibits greater morphological diversity than the rest of the genus." - Der genetische Artbegriff hat mit dem vorliegenden Beispiel keinerlei Schwierigkeiten. Selbstverständlich gehören auch die unterschiedlichsten miteinander fertilen Formen zur selben Art und darüber hinaus gehören auch sämtliche durch sekundäre Sterilitätsbarrieren voneinander getrennten, aber morphologisch weniger unterschiedlichen Formen aller Kontinente zur selben Art, und das bedeutet sehr wahrscheinlich, dass sich die ganze Gattung mit ihren mehr als 240 'Arten' genetisch auf eine Art reduziert.
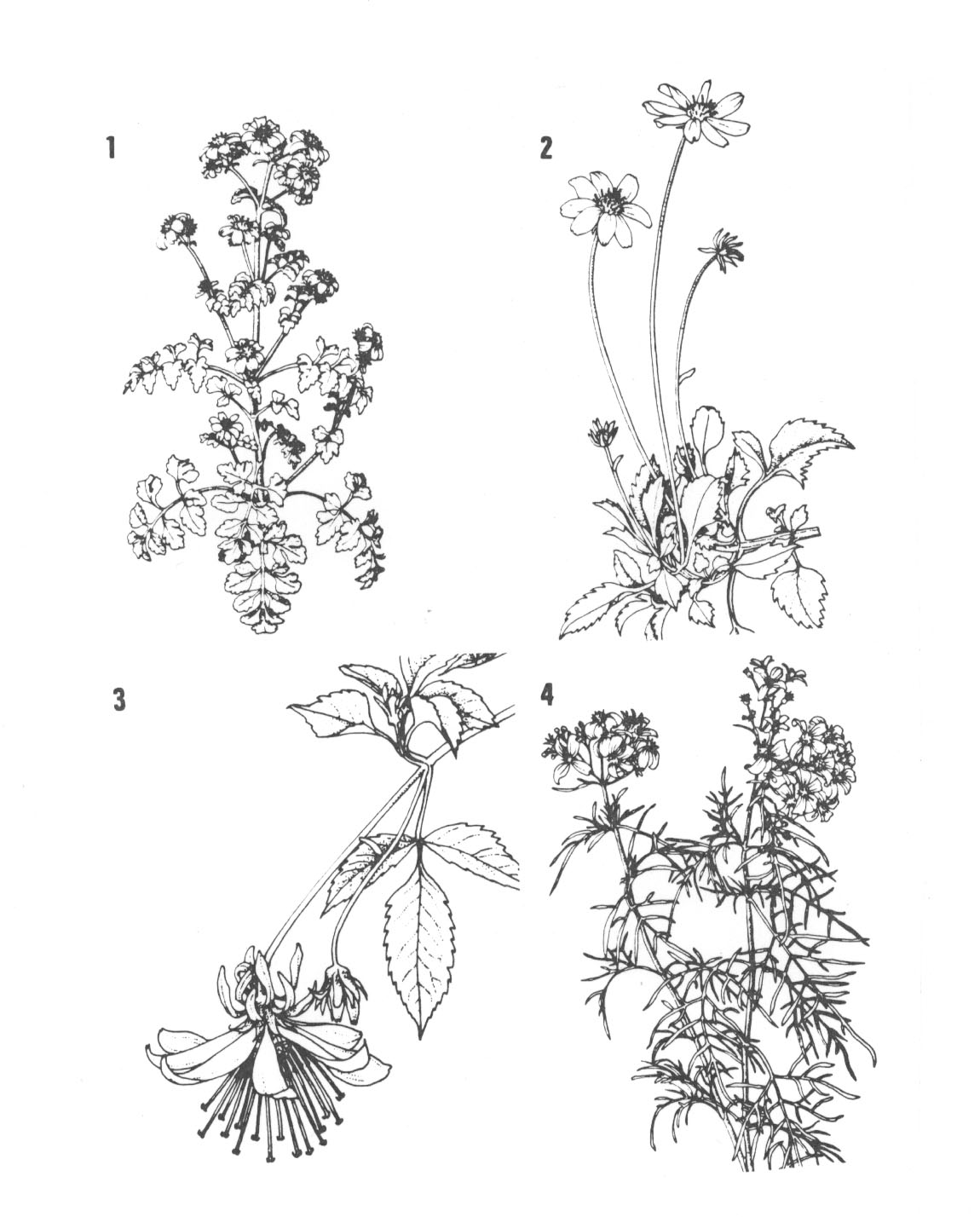
Die folgenden Abbildungen geben einige charakteristische Unterschiede zwischen hawaiischen Bidens-'Arten' wieder. Solche starken morphologischen Unterschiede halten viele Autoren davon ab, die miteinander fertilen Formen zur gleichen Art zu rechnen. Vergleicht man solche Unterschiede hingegen mit den oft noch stärkeren morphologisch-anatomischen Divergenzen zwischen Kulturpflanzen und ihren Ausgangsformen sowie den Unterschieden zwischen den Kultursorten selbst, so dürfte das Problem an Härte verlieren: Wenn wir bei den oft viel größeren Unterschieden keine Schwierigkeiten sehen, die Formen zur selben Art zu rechnen, wieso sollte das dann bei den meist geringeren Unterschieden der Wildformen der Fall sein? Wo immer Arten mehr Freiheit zur Realisation ihres genetischen Potentials sowie zur Degeneration haben (ob in menschlicher Obhut oder auf abgelegenen Inseln), verstärken sich auch die morphologischen, physiologischen, ökologischen etc. Unterschiede. Der Eliminierungsprozess aberranter Formen ist auf ozeanischen Inseln häufig geringer als auf den Kontinenten - oder in neodarwinistischer Terminologie: der 'Selektionsdruck' ist (zumindest) anfangs nicht so stark.
Nach Abbildung verschiedener Bidens-'Arten' geben wir die hauptsächlichsten Unterschiede zwischen den Kohlsorten wieder (nächste Seiten). Der Vergleich der Unterschiede der miteinander fertilen Bidens-Arten mit den morphologischen Unterschieden zwischen Wild- und Kulturformen sowie der Divergenz innerhalb der Kulturformen der verschiedenen Kohlsorten kann zur Klärung des Artbegriffs beitragen: Wildkohl, Blattkohl, Weiß- und Rotkohl, Wirsing- und Blumenkohl und viele andere Sorten werden nicht in den Rang eigener Kohlarten erhoben. Sowenig es sich hier um eigene Spezies handelt, genauso wenig handelt es sich bei den miteinander fertile Hybriden bildende Wildformen von Bidens um eigene Arten.
Abb. 40.1-4: 1. Bidens hillebrandiana ssp. polycephala from Maui. 2. Bidens mauiensis from Maui. 3. Bidens cosmoides from Maui. 4. Bidens menziesii ssp. filiformis from Hawaii. Aus Ganders und Nagata (1984).

Abb. 41: Morphologische Unterschiede beim Kohl: A Wild-Kohl (Brassica oleracea var. oleracea und verwandte Sippen, B Blattkohl (var. viridis), C Kohlrabi (var. gongylodes), D Rosen- oder Sprossenkohl (var. gemmifera), Weiß- bzw. Rotkraut oder Weiß- bzw. Rotkohl (var. capitata), F Wirsing (var. sabauda), G Blumenkohl oder Karfiol (var. botrytis). Aus Ehrendorfer (1978) nach Transeau, Sampson & Tiffany.
Was das Pflanzenreich generell betrifft, so finden wir hier noch häufiger als im Tierreich, dass "morphologisch sehr unterschiedliche Formen und sehr stabile Populationen noch Fortpflanzungsgemeinschaften bilden können" (Willmann 1985, p. 51). Der letztere Autor kommentiert den Versuch einiger Forscher, das Biospezies-Konzept (damit meint Willmann "die endgültige Abschottung" gegenüber anderen Populationen, "die absolute Fortpflanzungsisolation" als hauptsächlichstes Arttrennungskriterium [siehe jedoch Einschränkungen pp. 303 - 312]) für solche stark polymorphen Arten fallenzulassen, folgendermaßen (1985, pp. 51-52):
Grant (1977: 170) nimmt diese Vielfalt zum Anlaß einer anders ausgerichteten Kritik am Artkonzept. Er sieht die Unterscheidung von Arten und Unterarten nicht als ausreichend an, um die Situation in der Natur zu erfassen. Mehrere Rassen bilden für Grant zwar eine Art; außerdem aber möchte er (wie schon 1957 oder 1963: 340) weitere artähnliche Taxa unterscheiden können. So will er Populationen, zwischen denen nur geringer Genaustausch erfolgt ("Semispezies") und die somit zwischen dem Niveau der Rasse und dem der Art stehen, zu einer Einheit zusammenschließen, die umfassender als die Art sei: Mehrere allopatrische (und deswegen nicht in Genaustausch stehende) Semispezies bilden nach ihm eine Superspezies (Begriff von Mayr 1931), mehrere sympatrische oder teilweise sympatrische Semispezies ein Syngameon (Begriff von Lotsy 1925).21 Somit sei die Art im Sinne des Biospezies-Konzeptes nicht immer die umfassendste Fortpflanzungsgemeinschaft.
Dazu ist zu bemerken, daß es subjektiven Kriterien unterliegt, wann wir eine Population noch als Rasse, wann bereits als Semispezies im obigen Sinne ansprechen - oder gar als Art, die mit einer anderen Art in Genaustausch steht ("Art" im Sinne von Grant, nicht im Sinne des konsequenten Biospezies-Konzeptes). Von dieser Einschätzung hängt aber ab, ob eine Gruppe von Populationen eine Spezies oder eine Superspezies bzw. ein Syngameon darstellt.
Entscheidend aber ist der folgende Einwand. Mehrere Semispezies (oder "Spezies", zwischen denen begrenzter Genaustausch stattfindet) unter Umgehung des Zustandes der Biospezies als Superspezies oder Syngameon zusammenzufassen (so verstehe ich auch Grants Ausführungen 1957: 68), ist eine Unmöglichkeit, weil die Kriterien für die Biospezies (reproduktive Isolation zwischen Fortpflanzungsgemeinschaften) nicht umgangen werden können. Das merkt auch Grant (1977; Tafel 18.2), denn er führt die Kriterien der Art auch bei der Superspezies und dem Syngameon an. Er zieht daraus aber nicht den Schluß, daß diese beiden Begriffe dasselbe bezeichnen wie der Terminus "Biospezies".
Entgegen der Ansicht von Grant (oder auch Burger 1975: 48-50) gilt das Biospezies-Konzept unabhängig davon, wie umfassend und wie strukturiert "groups of interbreeding natural populations that are reproductively isolated from other such groups" sind. "A species is the most inclusive Mendelian population" (Dobzhansky 1970: 357), und nichts sonst. Ebenso wie der Begriff "Semispezies" sind auch die Termini Superspezies und Syngameon überflüssig (s. auch Key 1981: 441).
Hier wird der genetische Artbegriff konsequent angewandt und man kann Willmann aus dieser Sicht zu den obigen Ausführungen nur zustimmen.
"Bildung ist das Bewußtsein der Zusammenhänge" - hat A. Kaestner einmal festgestellt. Hier liegt einer der Stärken des genetischen Artbegriffs: Durch ihn wird die Aufdeckung oft ganz neuer, vorher vielleicht nicht einmal geahnter, unmittelbarer Zusammenhänge möglich. Ganz erstaunlich unterschiedliche Lebensformen, die als eigene Arten, Gattungen (vielleicht sogar als Familien) unterschieden wurden, können zur selben Mendelpopulation gehören. Die realgenetische Verwandtschaft steht nach Erweis der Tatsache, dass bestimmte, unterschiedliche Lebensformen "bei gegenseitigen Kreuzungen in allen Merkmalen den MENDEL'schen Gesetzen folgen", unmittelbar vor Augen.
SCHWÄCHEN:
Die Hauptschwäche des genetischen Artbegriffs liegt in der häufigen Nicht-Umkehrbarkeit seiner Grundaussage. Wenn unterschiedliche Lebensformen bei reziproken Kreuzungen in allen Merkmalen den Mendel'schen Regeln folgen, handelt es sich um Angehörige derselben Art. Der Umkehrschluss, dass Lebensformen, die in der Kreuzungsanalyse wegen unterschiedlicher Sterilitätsbarrieren nicht den Mendel'schen Regeln folgen können, deswegen zu zwei verschiedenen Arten gehören, ist jedoch nicht konsequent durchführbar. Ich habe das in allen Einzelheiten bei der Behandlung der postzygotischen Isolationsmechanismen aufgeführt und diskutiert. Sekundäre Sterilitätsbarrieren können ihre Ursache z.B. in genischer Degeneration (vgl. pp. 122-137), Chromosomen- und Genommutationen (pp. 145-168), Rickettsien- und Virus-Befall des Plasmas (pp. 129, 245/246) haben. Solche sekundären Sterilitätsbarrieren sind zwar z.T. reversibel, aber in vielen Fällen sind sowohl morphologisch gleichartige als auch recht unterschiedliche Lebensformen durch diese Mechanismen - einzeln oder in Kombination - so stark voneinander isoliert, dass der Erweis des unmittelbaren realgenetischen Zusammenhangs mit den größten Schwierigkeiten verbunden ist.
Es wäre ja auch zu schön, wenn wir mit dem genetischen Artbegriff nicht nur eine Fülle neuer Zusammenhänge, sondern zugleich ein absolutes Artabgrenzungskriterium in der Hand hätten. Die Sterilitätsbarriere als absolute Arttrennungsdefinition angewandt würde über die oben im Detail diskutierten Schwierigkeiten hinaus auch noch zu so merkwürdigen Schlussfolgerungen führen, wie die Artabgrenzung beim Menschen in sterilen Ehen (die Ehepartner gehörten - durch die Sterilitätsbarriere "erwiesen" zwei verschiedenen Arten an). [The risk of childlessness ... rises from 6% for 20 to 24 years old (women) to 9% for those 25 to 29, to 15% for women who marry in their early 30's, and then increases much more sharply." (J. Menken, J. Trussel und U. Larsen 1986, p. 1389: Age and infertility; Science 233, 1389-1394.)]
Kein vernünftiger Biologe würde auf diese Weise beim Menschen Arten voneinander trennen. Pathologische Veränderungen, die zur Sterilität führen, müssen grundsätzlich vom genetischen Artbegriff ausgeklammert werden. Wichtig ist jedoch dabei festzuhalten, dass pathologisch-genische Veränderungen nicht nur einzelne Individuen, sondern auch Populationen von Individuen betreffen können, wobei dann verschiedene Populationen mit unterschiedlichen genischen Abbauprozessen (die innerhalb der Populationen noch verkraftet werden) miteinander eine Sterilitätsbarriere aufweisen können.
Dass der Umkehrschluss wegen sekundärer Sterilitätsbarrieren nicht konsequent möglich ist, das sahen wohl auch schon die großen klassischen Genetiker, die sich für den genetischen Artbegriff eingesetzt haben. Bei der Behandlung des Hieracium-Beispiels von Nilsson ist uns der Fall (p. 63) bereits begegnet. Viele Hieracium-'Arten' pflanzen sich durch Agamospermie (Embryobildung aus diploiden und unbefruchteten Eizellen) fort. Wir finden hier also asexuelle Vermehrung. "Wie diese zustande kommt, ist in diesem Zusammenhang von sekundärer Wichtigkeit. Sie mag Apomeiose, Aposporie, Nuzellarembryonie sein. In jedem Falle ist die Vermehrung ganz vegetativ, als ob sie durch eine Bulbille oder Stolone geschehen würde. Die Biotypen sind in diesem Falle wie bei der Autogamie konstant, aber vegetativ konstant." Wie wir auf den Seiten 63-66 gesehen haben, lehnt Nilsson die uferlose Speziesmacherei bei Hieracium und anderen Arten gut begründet ab. Der Umkehrschluss, dass die nicht miteinander kreuzbaren Hieracium-'Arten' tatsächlich 'echte' Arten sind, ist in diesem und vielen weiteren Fällen wegen der sekundären Sterilitätsbarrieren nicht möglich. Nilsson versucht dennoch an verschiedenen Stellen seines Werkes die Artdefinition im ursprünglich genetischen Sinne zu retten. So schreibt er z.B. p. 351:
Man muss daran unentwegt festhalten, dass bei der Spezies die Amphimixis der zusammenhaltende Moment, die Inkompatibilität und die Avitalität, beide ebenso wichtig, der differenzierende Moment ist. Falls der Begriff Spezies überhaupt eine Meinung haben soll, kann in dieser Formulierung keine Auflockerung erlaubt werden. [Alle Hervorhebungen im Schriftbild von Nilsson.]
- Der Autor steht jedoch vor der unausweichlichen Tatsache, dass durch sekundäre Sterilität bei zahlreichen Arten (Hieracium, Taraxacum, Alchemilla, Rosa, Rubus etc. werden von Nilsson selbst behandelt) die Artdefinition durch den Umkehrschluss nicht durchführbar ist. Für solche Fälle schreibt er p. 284:
Für die konsequente Fassung und Entfaltung des Speziesproblems ist...ein Ausgehen von den Allogamen eine unbedingte Notwendigkeit. Und dieses nicht nur deshalb, weil sie eine Präponderanz aufweisen, die fast zur Unbedeutendheit der anderen Gruppen führt, sondern auch aus dem Grunde, dass sie das normale genische Rekombinationsspiel darbieten. Von dem Normalen auszugehen, um das Abweichende, das Verwickelte zu deuten, ist ja ein gewöhnliches, gesundes wissenschaftliches Prinzip. Ein umgekehrtes Vorgehen...kann nur zu falschen Schlußfolgerungen führen. [Alle Hervorhebungen im Schriftbild von Nilsson.]
Wenn man die normalen (und nicht die durch sekundäre Sterilitätsbarrieren voneinander getrennten) Formen zum Maßstab für den Artbegriff macht, könnte man vielleicht den absoluten genetischen Artbegriff (d.h. mit Umkehrschluss) beibehalten. Auf der anderen Seite ist die Problematik Hunderttausender Zwillingsarten (nach White sogar Millionen) und wohl noch größerer Zahlen morphologisch unterscheidbarer durch sekundäre Sterilitätsbarrieren voneinander abgrenzbarer weiterer 'Arten' unübersehbar. Nilssons Hinweis von dem geringen Umfang solcher Gruppen ist beim heutigen Kenntnisstand nicht mehr haltbar.
Da die Verabsolutierung der Sterilitätsbarriere als artentrennendes Kriterium in die ganze Fülle der oben an zahlreichen konkreten Beispielen diskutierten Schwierigkeiten und Widersprüchen führen würde, bleibt dem genetischen Artbegriff gar nichts weiter übrig als zwischen primären und sekundären Sterilitätsbarrieren zu unterscheiden und nicht undifferenziert den Umkehrschluss zum absoluten Maßstab für den Artbegriff zu machen.
Wohin überdies der undifferenzierte Umkehrschluss in der Wissenschaft führen kann, sei an einem aktuellen Beispiel verdeutlicht:
H. von Ditfurth kommentiert (1984, p. 162) Erbens Einwände zu den wissenschaftstheoretischen Überlegungen Poppers:
Die Art und Weise zum Beispiel, in der dieser Philosoph das berühmte Induktionsproblem abgehandelt hat, hält Erben für wenig gelungen. Und das nicht minder berühmte "Falsifikations-Kriterium"? Die definitorische Orientierungshilfe, nach der eine Hypothese nur dann wissenschaftlichen Charakter beanspruchen kann, wenn sie nachprüfbar und wenigstens im Prinzip widerlegbar ("falsifizierbar") ist?...Erben hält auch diesen Popperschen Einfall nicht für sehr glücklich. Im Gegenteil, er hält das Falsifikationsprinzip für den Ausdruck einer "Konzeptionsschwäche" innerhalb des Systems der Popperschen Philosophie. Begründung: Wenn es konsequent angewandt werde, könne es zu unsinnigen Urteilen führen. Erbens Beispiel: "Die Astrologie vertritt zwar Nonsens, aber sie ist falsifizierbar, also müßte sie theoretisch als legitim wissenschaftlich klassifiziert werdenl!"*
Das ist nun wahrlich starker Tobak. Muß man Erben, der nicht müde wird, seinen zahllosen Kontrahenten logische Mängel, "unbedachte Schnelldenkerei" und Schlimmeres vorzuwerfen, muß man ihn wirklich erst darauf hinweisen, daß er hier "notwendige" und "hinreichende" Bedingungen durcheinanderbringt und daher bei einem falschen Umkehrschluß landet? Zwar kann eine These, die grundsätzlich nicht widerlegbar ist, nicht als wissenschaftliche These gelten. Aber deswegen ist doch nicht, umgekehrt, jeder widerlegbare Unsinn schon Wissenschaft. Wenn ich die Behauptung aufstellen würde, der Mond bestehe aus erstklassigem Edamer Käse, dann ließe sich das mühelos "falsifizieren". Aber daraus würde sich doch nun auf keinem logischen Wege die Folgerung ableiten lassen, daß der Ausgangsbehauptung wissenschaftlicher Rang zukäme.
[*T. Theocharis und M. Psimopoulos machen in ihrem ansonsten recht lesenswerten Beitrag WHERE SCIENCE HAS GONE WRONG (Nature 329, 595-598; 1987) einen ähnlichen Fehler, ebenfalls einen falschen Umkehrschluss, wenn sie über Poppers Ansatz p. 595 schreiben: "The criterion of the scientific status of a theory is its falsifiability". (Example: "The earth is (approximately) a sphere" is not a scientific statement because it is not falsifiable; whereas "The Earth is a flat disk" is indeed scientific.) - worauf R. Facius (Nature 330, p. 689) richtig stellt: ''Popper's demand of falsifiability requires a theory (as opposed to a statement of a singular empirical fact) to have among its logical deductions at least one statement concerning a new, single, empirical fact (...), which in principle can be contradicted by a statement about an empirical observation or the result of an experiment."]
Das Prinzip lässt sich (mit positivem Vorzeichen) auch auf unsere Frage nach der genetischen Artdefinition anwenden: Zwar gehören alle kreuzbaren Formen mit fertiler Nachkommenschaft zur selben Art, aber deswegen gehören doch nicht alle Formen mit x-beliebiger (und z.T. reversibler) Fertilitätseinbuße schon verschiedenen Arten an (vgl. pp. 122 - 195).
Dem genetischen Artbegriff kommt das Verdienst zu, eine Fülle vorher nicht geahnter, direkter realgenetischer Zusammenhänge aufgedeckt zu haben und weiter aufzudecken.
Was den Ursprung der Arten betrifft, gehören die Befürworter des genetischen Artbegriffs häufig zu den Kritikern der Synthetischen Evolutionstheorie. Das ist wohl auch ganz unvermeidlich, denn der Neodarwinismus zieht die Artgrenzen häufig bereits zwischen Formen, "welche bei gegenseitigen Kreuzungen in allen Merkmalen den MENDEL'schen Gesetzen folgen..." (de Vries). An dieser Stelle sei an weitere Wegbereiter der Genetik erinnert. Nach Bateson waren die Fragen nach dem Ursprung der Arten nach der 'Wiederentdeckung' der Mendel'schen Regeln wieder völlig offen (vgl. Zitate oben). 1924/1928 schrieb er p. 406 :
Mendelian analysis led to the discovery of the transferable characters, not merely in sporadic instances but as a group, and the study of their behaviour enabled us to avoid endless misinterpretations into which our predecessors had consistently fallen. If we now have to recognize that the transferable characters do not culminate in specific distinctions, the acknowledgement will not come from us alone. The old belief of systematists that real species differ from each other in some way not attainable by summation of varietal characters is no longer contestable, and we know now upon what to concentrate.
Auf Seite 407 bemerkt er: "Perhaps the true specific characters belong to the cytoplasm, but these are only idle speculations."
1913 [Nachdruck 1979], p. 236/237 liefert der Autor einige wichtige Punkte zur genetischen Artdefinition:
Complete fertility of the results of inter-crossing is and I think must rightly be regarded as inconsistent with actual specific differences.
Zum Umkehrschluss stellt er im selben Werk fest (p. 247):
Sterility as a result of crossing, as well as that which is alleged sometimes to arise in consequence of changed conditions, is at best a negative characteristic and there are endless opportunities for mistake and interpretation in studying features of this kind.
Die Bedeutung der Selektionstheorie für die Frage nach dem Ursprung der Arten wurde von de Vries, Johannsen, Bateson u.v.a. nachdrücklich in Frage gestellt. So sei noch einmal Bateson aus seinem Werk von 1913 zitiert, in dem er an zahlreichen Stellen auf den Selektionsgedanken eingeht. Auf der Seite 134 schreibt er z.B.:
...the stringency of Selection is frequently so far relaxed that two or more equally definite forms of the same species can persist side by side. There is no doubt that this is the simple truth, but when once that truth is perceived it is useless to invoke the control of Selection as the factor to which definiteness of type in general must be referred.
An zahlreichen Beispielen wird in diesem Werk die Selektionstheorie in Frage gestellt. Ich kann darauf an dieser Stelle nicht weiter eingehen und möchte den Leser daher auf diese Arbeit Batesons verweisen. Im Gegensatz zu dem oben zitierten 'alten Glauben' der Systematiker, sah Bateson natürlich die Differenz zur systematischen Praxis und schrieb bereits 1909/1914, p. 286, sehr treffend:
Es steht erstens einmal fest, daß die Spaltung in zahllosen Fällen eine Rolle spielt bei der Bildung und Erhaltung von Eigenschaften, welche von Systematikern als ein diagnostisches Merkmal für Arten angesehen werden.
(p.287:) Es braucht kaum betont zu werden, daß eine große Zahl der Eigenschaften, von denen man jetzt weiß, daß sie spalten, imstande wären, in den Augen der meisten Systematiker spezifische Unterschiede darzustellen, wenn die betreffenden Tiere oder Pflanzen ihnen von Sammlern gebracht würden. Wir können sogar sicher sein, daß eine große Zahl ausgezeichneter Spezies, die von Entomologen und Ornithologen beispielsweise allgemein anerkannt sind, sich bei Kreuzungsversuchen sofort als analytische Varietäten erweisen würden, die sich lediglich durch die Anwesenheit oder Abwesenheit bestimmter Faktoren voneinander unterscheiden.
Zur Frage nach dem Ursprung der Arten heißt es in demselben Werk (p. 291):
Die Anschauung, daß die Evolution durch allmähliche Umformung von Massen von Individuen infolge der Summation unmerklicher Veränderungen zustandekommt, erweist sich durch das Studium der Genetik sofort als falsch.
Interessanterweise hatte Bateson schon 1909 ein ganzes Forschungsprogramm zum Artbegriff konzipiert, dessen Durchführung heute noch in wesentlichen Punkten am Anfang steht. Zum Thema 'mögliche Grenzen für Neukombinationen' hebt er (p. 72) hervor:
Die Bestimmung dieser Grenzen ist eine der wichtigsten Aufgaben, die unser noch harrt.
(p. 73:) Zweitens müssen wir annehmen, daß diese vererbbaren Eigenschaften an eine gewisse Grundveranlagung gebunden sind, und die Fähigkeiten oder Kräfte, welche gemeinsam jene notwendige Grundlage bilden, lassen sich vielleicht von den übertragbaren Eigenschaften selbst trennen. Die Aufdeckung der Grenzen, welche der Übertragbarkeit der Eigenschaften gesetzt sind, wäre von größter Bedeutung und stände in engster Beziehung zu dem Problem vom letzten Wesen der Arten.
Genau an dieser Stelle wird das Problem der primären Artbarriere aufgeworfen. Bevor wir auf diesen Punkt ausführlich im Zusammenhang mit Lamprechts Artbegriff zu sprechen kommen, möchte ich noch einmal die Differenz zwischen den Befürwortern des genetischen und des neodarwinistischen Artbegriffs zum Ursprung der Arten hervorheben. Der Genetiker Wartenberg schreibt 1967, p. 165:
Nicht alle Forscher der Genetik sahen in ihrer Arbeit eine Artbildungsphysiologie. Im Gegenteil: Es gab unter den Genetikern von Weltruf entschiedene Gegner der Evolutionstheorie. Vor allem waren es die Engländer Galton und Bateson, der Däne Johannsen und einige Forscher aus der schwedischen Genetikerfamilie Nilsson, dessen Buch "Synthetische Artbildung" von 1953 ein mit viel gründlichem Wissen geführter Kampf gegen die Evolutionstheorie ist.
Und auf der Seite 176 fasst er seine Studie wie folgt zusammen:
Die Frage, ob die Genetik Beweise für die Thesen der Evolutionstheorie geben kann, muß mit einem Nein beantwortet werden.