DETAILS ZU DEN INTERSPEZIFISCHEN GENEN
Sehen wir uns die Lamprechtschen Gedankengänge zum Begriffspaar der inter- und intraspezifischen Gene noch etwas näher an (1974, pp. 366/367):
Zusammenfassend können die interspezifischen Gene folgendermaßen gekennzeichnet werden:
1. Die Allele eines interspezifischen Gens sind immer auf verschiedene Spezies oder auch höhere Kategorien verteilt. Sie sind demnach Arten bzw. höhere Kategorien trennend.
2. Solche Allele bedingen die unüberbrückbare Barriere zwischen naturbedingten Spezies und höheren Kategorien.
3. Jedem arteigenen Allel eines interspezifischen Gens für die Gestaltung des vegetativen Teils der Pflanze entspricht ein auf dieses abgestimmte Allel eines interspezifischen Gens für den floralen Teil. Die Aufgabe des letzteren Allels besteht in der Umbildung vegetativer Teile in funktionstaugliche florale. Diese aufeinander eingestellten Allele bzw. Gruppen von Allelen interspezifischer Gene bilden das sog. vegetativ-florale Genensystem. Jede Veränderung irgendeines der diesem System angehörigen Allele führt zu vollkommener Sterilität.
4. Die artfremden Allele von interspezifischen Genen mutieren im somatischen Gewebe der sich entwickelnden Pflanze stets zum arteigenen Allel zurück. Je nachdem, in welchem Stadium der Entwicklung dies stattfindet, tritt dann die Manifestation des arteigenen Allels auch visuell erkennbar zutage und führt in vielen Fällen am Gipfel der Pflanze zu Hülsen- und Samenbildung.
Die intraspezifischen Gene verhalten sich, wie aus oben Angeführtem hervorgeht, hinsichtlich jeder der in den vier Punkten erwähnten Erscheinungen grundverschieden. Die Allele intraspezifischer Gene sind nicht auf verschiedene Arten verteilt, sondern binnen diesen beliebig miteinander kombinierbar, wenn duch das hierbei erzielte Ergebnis höchst verschieden ausfallen kann. Außer sehr vitalen Formen können physiologisch schwache, gegen Parasiten anfällige und auch letale Formen auftreten. Die enorme Kombinationsmöglichkeit von intraspezifischen Genen liegt der Entstehung der zahlreichen Rassen zugrunde, die sowohl in der Natur wie im Kreuzungsexperiment auftreten.
Intraspezifische Gene können sowohl im vegetativen wie im floralen Teil der Pflanze unabhängig wechselnde Merkmale bedingen. Sie können sowohl in dieser wie auch in anderer Hinsicht pleiotrope Wirkungen an den Tag legen. So gibt es z. B. bei Pisum eine Anzahl von Genen, die den Längen-Breiten-Index von sowohl Stipel, Blättchen wie auch Kelchblättern pleiotrop beeinflussen (s. HÄRSTEDT 1950 u. o.). Es sind dies Fälle von typischer Pleiotropie, indem jedes einzelne dieser Gene mehr als ein Merkmal eines Organismus in gewisser Richtung beeinflußt.
Auf den Seiten 561/562 folgen Beispiele für interspezifische Gene:
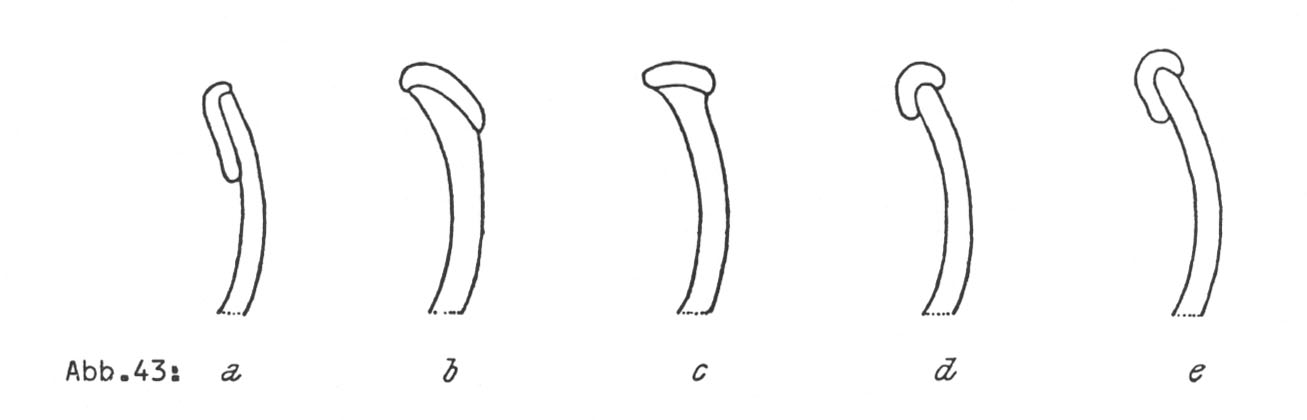
Zwei in ihren Einzelheiten in sehr großem Umfang studierte Fälle seien hier erwähnt (Näheres bei L 1966). Der erste bezieht sich auf die beiden in Süd- und Zentralamerika heimischen Phaseolus-Arten vulgaris und coccineus, die durch die Allele von zwei interspezifischen Genen getrennt sind. Das vulgaris-Allel des einen dieser Gene, i-Int, bedingt das Herablaufen der Narbe auf der Innenseite der Griffelspirale, das entsprechende Allel von coccineus, i-int hat das
Herablaufen der Narbe auf der Außenseite der Griffelspirale zur Folge. Die Allele des zweiten interspezifischen Gens, i-Epi (von vulgaris) und i-epi (von coccineus) sind für die epi- bzw. hypogäische Stellung der Keimblätter am Stamm verantwortlich. Die Abbildungen 199 und 200 zeigen diese Verhältnisse.
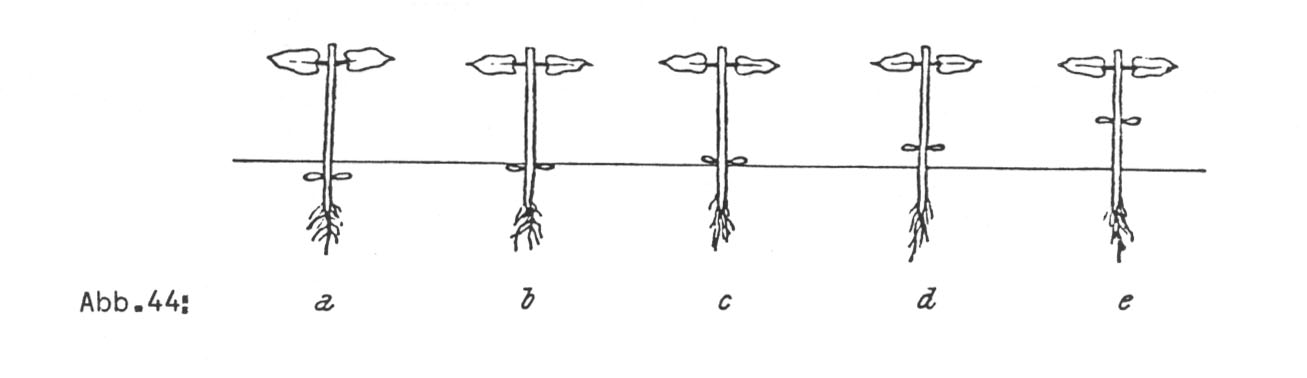
Die Kreuzungen wurden sowohl mit Kultur- wie mit Wildmaterial in beiden Richtungen ausgeführt. Das Ergebnis war dasselbe. In bezug auf die beiden artentrennenden Merkmale waren die Bastarde intermediär. Man vergleiche die Abbildungen 43 und 44.
Mit vulgaris als Mutter zeigte die F2 folgende Spaltung:
1 vulgaris-Narbe: 2 Bastard-Narbe: 0 coccineus-Narbe
Mit coccineus als Mutter war die Spaltung wie folgt:
1 coccineus-Narbe: 2 Bastard-Narbe: 0 vulgaris-Narbe
In vollkommen entsprechender Weise spaltete die Lage der Kotyledonen am Stamm, nämlich mit vulgaris als Mutter:
1 epigäisch: 2 intermediär: 0 hypogäisch
bzw. mit coccineus als Mutter:
1 hypogäisch: 2 intermediär: 0 epigäisch
Eine fertile Pflanze mit Homozygotie in den artfremden Allelen, d. h. mit einem für die Spezies fremden Merkmal, konnte in den zahlreichen, etwa eine halbe Million Individuen umfassenden Material während 35 Jahren niemals beobachtet werdert.
Als zweites Beispiel folgt die Kreuzung Chrysanthemum carinatum x Chr. coronarium und reziprok, die ein im Prinzip gleiches Verhalten zeigt. Weiter sei hervorgehoben, dass nach Lamprechts Meinung artfremde Allele vom Plasma nicht nur in der Ontogenese nicht homozygot exprimiert, sondern während der Keimzellbildung (falls es soweit überhaupt kommt) auch nicht redupliziert, nicht "erneuert" werden. Warum nicht? Nach Lamprecht ist das Plasma auch für die Erneuerung von "Progenen" zuständig, die nun ihrerseits für die Synthese (= Replikation) der Gene während der Keimzellbildung notwendig sind. "Die Progene werden nur einmal je Generation, bei der Reduktionsteilung, erneuert. Ihre Gesamtheit wird als Progenom bezeichnet" (p. 366). Er behauptet, dass 'jedem Genallel ein auf dasselbe eingestellte Progen zugeordnet ist'. "Die Progene liegen gleichwie die Gene in den Chromosomen" (p. 366). Daraus folgt, dass artfremde Allele bei der Keimzellbildung auch nicht redupliziert werden können, wohl aber in den Mitosen der von ihm untersuchten Hybriden, da sie ja die "Progene" noch in dem väterlichen Chromosomensatz vorzuliegen haben.
Die experimentellen Befunde an Phaseolus sind inzwischen wiederholt bestätigt worden (Smartt 1970, Haq et al. 1980, Sarbhoy 1980, Shii et al. 1982), aber das Progenkonzept ist eine Hypothese aus vormolekularer Zeit, die in dieser Form mit vielen Schwierigkeiten behaftet ist. Bereits 1944 war diese Hypothese im Detail veröffentlicht worden. Lamprecht schreibt 1944, p. 103:
Die Gene, d. h. Allele, werden von Mitose zu Mitose neugebildet, vermehrt. Wir können uns diesen synthetischen Prozess so vorstellen, daß es Stoffe enzymähnlicher Natur gibt, die für diese Gensynthese aus ihren unmittelbaren Vorstadien verantwortlich sind. Hierbei sind die Gene stets als in einer bestimmten Genform vorhanden aufzufassen. Diese die Gensynthese besorgenden Stoffe, wir können sie Progene nennen, da sie für die Gene da sind, gehören in Übereinstimmung mit den Beobachtungen jener biologisch zentralen Gruppe von chemischen Verbindungen an, die beim Generationswechsel erneuert werden. Für ihre Erneuerung ist wahrscheinlich der Nukleolus verantwortlich. Zu dieser Gruppe werden auch jene Stoffe gehören, die mit dem Altern der Organismen schwinden und beim Generationswechsel erneuert werden. Und diese Erneuerung der Progene von Generation zu Generation scheint für die wirklich arttrennenden Gene (Allele) nur im arteigenen Plasma möglich zu sein. Man kann sich hier fragen, wo sich die Progene befinden, in oder außerhalb der Chromosomensubstanz. Die Tatsache, daß die Elimination von wirklich arttrennenden Genen beide Geschlechter gleich trifft sowie mit Hinblick auf den Umstand, daß die männlichen Gameten kein oder nur höchst unbedeutende, gewöhnlich nicht feststellbare Spuren von Plasma erhalten, spricht stark dafür, daß die Progene (und vielleicht auch die letzten Vorstadien der Gene) der Chromosomensubstanz angehören. Die Progene, wie auch andere, oben angedeutete biologisch zentral stehende Stoffe, werden meiner Ansicht nach beim Generationswechsel durch die Wirkung des Nukleolus erneuert. Und diese Erneuerung scheint in übereinstimmung mit den Erscheinungen in Kreuzungen von Ph. vulgaris x multiflorus...in artfremden Plasma zu unterbleiben.
Zu den oben zitierten nicht überführbaren Genen bei den Bohnenarten schreibt er (p. 120):
Die Mutation von Hyp zu Epi und von Ext zu Int mit einer Frequenz von wenigstens 5 - 6 %, bei alleiniger Berücksichtigung der sich zu Pflanzen entwicklenden Zygoten sogar 86 %, beweist, daß es sich hier nur um zwei verschiedene Allele desselben Gens handeln kann. Aus der Erscheinung, daß diese Gene wohl somatisch gut nebeneinander reproduziert werden können nach der Bildung der Tetraden aber unmittelbar in sehr großem Prozent zu Zellendegeneration führen, kann folgendes geschlossen werden: für die Synthese der Gene sind Stoffe verantwortlich, von mir als Progene bezeichnet, die während jeder Generation nur einmal, und zwar bei der Reduktionsteilung erneuert werden.
(p. 121:) Die Progene gehören offenbar derselben Gruppe von biologisch nentralen Stoffen an, die für das Altern verantwortlich sind und auch beim Generationswechsel erneuert werden. Ihre Erneuerung wird wahrscheinlich durch die Wirkung des Nukleolus besorgt. In den wenigen Fällen, wo Pflanzen mit den Genen Hyp und Ext ebensolche Nachkommen in einer oder maximal bis drei Generationen gaben, beruht dies wahrscheinlich darauf, daß ein wenig von ihren Progenen von der vorherigen Generation hat weitergegeben werden können.
(p. 124:) Das Plasma liefert das letzte Vorstadium des Gens (das vielleicht schon ein, aber inaktives, Allel des betreffenden Gens darstellt) und dieses wird unter dem gleichzeitigen Einfluß von Progen und Gen in das Gen umgewandelt, wobei sich dies stets auf eine bestimmte Genform (Allel) bezieht. Fehlt entweder das Gen oder Progen, so unterbleibt die Synthese der betreffenden Genform. Wie schon erwähnt ist anzunehmen, daß jede Art - mit Ausnahme für die primär arttrennenden Gene - Progene für sämtliche Allele ihrer Gene enthält.
Der Ansatz ist im Grunde genommen denkbar einfach: In den von ihm untersuchten Hybriden findet er für die nicht überführbaren Merkmale einen intermediären Erbgang, d.h. dass hier noch beide Allele repliziert werden und funktionieren (im Gegensatz zu den Befunden Ohnos bei den Forellen- und Wachtelhybriden, bei denen ein Teil des väterlichen Genoms erst gar nicht "angeschaltet" und ein weiterer Teil verspätet exprimiert wurde). Wenn beide Allele in den Hybriden funktionieren (repliziert und "angeschaltet" werden) - warum sind sie dann nicht auch reziprok überführbar? Warum erhält man dann nicht auch die Rekombinanten ?
Hier blockiert nach Lamprechts Meinung das Plasma, welches die artfremden Allele in der Meiose nicht reproduzieren kann (genauer gesagt, welches die 'Progene' für die Replikation der artfremden Allele nicht synthetisieren kann). Warum klappt dann aber noch die Replikation in allen Mitosen durch die ganze Ontogenese der Hybriden hindurch? Die "Progene" (noch vom Plasma der Elternart produziert) liegen in den Chromosomen. Die Erneuerung der Progene für die artfremden Allele unterbleibt in der Meiose (dirigiert vom arteigenen Plasma) und die artfremden Merkmale können nicht überführt werden.
Blixt kommentiert Lamprechts MONOGRAPHIE DER GATTUNG PISUM zu dem hier angeschnittenen Thema (1976, p. 61) wie folgt:
As might be expected, some space is devoted to Lamprechts own hypothesis on intra- and interspecific genes, but the emphasis is not excessive. This hypothesis, as well as some of his terminology, undoubtedly reflects the fact that Lamprecht worked in the age of classical genetics and that he belonged to a group who formed their views of the gene long before the "molecular" era and the discovery of the genetic code. This, however, in no way diminishes the value of his important work in gene-manifestation and linkage... Pea geneticists will not want to be without this book.
Der Einwand ist sicher richtig, erklärt jedoch nicht die empirisch gewonnenen Resultate (im übrigen stammt ja auch der ganze Mendelismus aus klassischer Zeit und ist in den letzten Jahrzehnten molekulargenetisch systematisch untermauert und bestätigt worden - spezifische Genkonzepte aus dieser Zeit haben sich jedoch in mehreren Fällen als unrichtig erwiesen, z. B. Goldschmidts Proteingen-Konzept).
Es könnte ja auch sein, dass Lamprecht mit seinem Plasmaregulationsansatz die einseitig DNA-orientierte molekulargenetische Epoche übersprungen hat und jetzt mitten in der molekular-entwicklungsgenetischen Epoche landet, in der die Kern-Plasma-Relation und die Regulationsprozesse über das Plasma in den Vordergrund der Forschung rücken (vgl. die obigen Zitate, in denen die Bedeutung des Plasmas für die Regulation hervorgehoben und begründet wird).
Natürlich kann sich manches Detail des Lamprechtschen Ansatzes durch die weitere Forschung noch als falsch erweisen. So ist z.B. seine immer wiederkehrende Behauptung, dass die 'Mutationen nicht die Gene selbst treffen, sondern stets die Progene' durch die umfangreichen Forschungen und Ergebnisse der Molekularbiologie klar als falsch erwiesen (zum Thema Mutationsprozesse vgl. man z.B. Auerbach 1976, Alberts et al. 1983 oder irgendeines der modernen molekularbiologischen Lehrbücher).
Inzwischen haben wir jedoch auch gelernt, dass Mutationen mittelbar ausgelöst werden können. So schreiben beispielsweise Anderson, Richardson und Davies in der Zusammenfassung ihres Artikels THE GENOTOXIC POTENTIAL OF BASES AND NUCLEOTIDES (1981, p. 265):
This study was conducted to determine if naturally occurring cell constituents could themselves cause mutation. All the bases and their corresponding nucleotides have been shown to produce chromosome damage in P 338 mouse lymphoma and chinese hamster ovary cells in culture. ...Such damage probably arises as a result of imbalanced DNA-precursor pools. Thus mutagenic events may arise by mechanisms unrelated to direct alterations of DNA.
De Serres hat zu diesem Thema 1985 ein ganzes Buch mit dem Titel GENETIC CONSEQUENCES OF NUCLEOTIDE POOL IMBALANCE herausgegeben.
Kubinski et al. stellen in ihrem Artikel DNA-CELL-BINDING (DCB) ASSAY FOR SUSPECTED CARCINOGENS AND MUTAGENS (1981, p. 96) fest:
The DCB assay is based on the observation that nucleic acids, including DNA, form complexes with proteins and with other nucleic acid molecules in the presence of active carcinogens.
...That proteins are the most likely target molecules in the cellular membrane has been shown by experiments in which DNA binding was abolished following a short exposure of cells or of purified membranes to proteolytic enzymes.
Hélias und Proust berichten 1982 über MUTATIONS INDUCED BY HORMONAL DROSOPHILA MELANOGASTER (p. 147): "...hormonal imbalance...disorganizes the synthesis of genetic information so that hereditary faults may appear in the individual's descendents."
Kilby schreibt über die Mutagenese (1983, p. 112):
Mutagenesis is a multi-step process which starts when the mutagen enters or is formed in the cell and interacts with the DNA. It ends with the production of a mutant individual. In between, a variety of cellular processes participate which include repair, DNA replication, RNA and protein synthesis. Monitoring can accordingly be performed at a variety of points which inevitably become less and less accurate as indicators of exposure as more of the components of cellular response to exposure are built into the endpoint measured.
Vgl. auch Nicoloff (1981), Kunkel et al. (1982) und Murthy und Prema (1983).
Aber auch Auerbach stellt (1976, pp. 349/350) fest:
During what I have called the fourth period of mutation research (Chapter 1), all efforts were directed towards analysing the action of those mutagens that react directly with DNA. Research on the indirectly acting ones...was put aside. The more recent realization that the production of mutations by almost any mutagenic treatment depends to a large extent on biochemical processes in the cell as a whole, opened the way for attempts at influencing mutation frequencies indirectly via chemicals that inhibit or deflect these processes. ( - Womit die obigen Zitate zur fünften und neuesten Forschungsperiode gehören.)
Doch auch bei diesem Thema muss einschränkend hinzugefügt werden, dass es nicht ausschließlich eine bestimmte Stoffklasse ist, durch die die indirekte Mutationsauslösung bewerkstelligt wird (nach Lamprecht waren es ja immer die für die Replikation notwendigen "Progene").
Immerhin kommt die indirekte Mutationsauslösung dem Lamprechtschen Ansatz näher, ohne ihn jedoch im Detail zu bestätigen.
Zurück zum Progen-Konzept als Vorraussetzung für die Replikation: Zunächst sei vermerkt, dass Dillon im Zusammenhang mit der Genfunktion auf der Transkriptionsebene (1983 - vgl. Zitat p. 219 ) von "pregenes" spricht, die jedoch mit dem Lamprechtschen Konzept nur insofern etwas zu tun haben, als sie als Vorstadien der voll funktionsfähigen Gene ebenfalls vom Plasma aus dirigiert werden sollen.
Auf der Transkriptionsebene haben wir uns ja inzwischen schon an den Gedanken einer Genhierarchie gewöhnt (Jacob-Monod-Modell mit Struktur-, Operator-und Regulatorgenen mit Repressor- und Aktivatorproteinen, aber es gibt noch zahlreiche offene Fragen der Genregulation bei Eukaryonten - hervorragende Zusammenfassung z.B. bei Alberts et al. 1983). Dass jedoch Regulatorgene oder selbst die vermuteten 'master regulatory genes' (Alberts et al.p. 444) während der Keimzellbildung ausschließlich durch spezifische vom arteigenen Plasma produzierte 'Stoffe' repliziert werden sollen - darüber ist bislang nichts bekannt. (Wir möchten jedoch noch einmal an die Befunde der entwicklungsphysiologischen Forschung erinnern, dass ein großer Teil der Genregulation während der Differenzierung vom Eiplasma ausgeht - vgl. Zitate oben. )
Auf der anderen Seite könnte man sich aufgrund der bisherigen Ergebnisse der molekular-biologischen und entwicklungsphysiologischen Forschungen ein übergreifendes Regulationssystem vorstellen, das auch mit der Replikation in der Meiose befasst ist - aber wir wissen derzeit zuwenig darüber. Jedenfalls lässt sich Lamprechts Progen-Konzept bislang mit keinem der molekular-biologischen Konzepte und Forschungsergebnisse eindeutig identifizieren, - obwohl man hier an Themen denken könnte, wie etwa an die (in der Meiose für Crossing over und Chiasmata-Bildung zuständigen) Recombination nodules im synaptischen Komplex oder an die Replikons als Replikationseinheiten (deren Zahl mit 20 000 bis 30 000 im haploiden Säugergenom und noch höheren Zahlen in frühontogenetischen Stadien - Literatur bei Alberts et al. 1983, Lewin 1985 - sich der Gesarntzahl der Gene nähert). Nach Ohno wurden die paternellen Allele vieler Genloci in seinen Hybriden ignoriert (vgl. p. 224), und weitere Befunde zeigen, dass bei Kerntransplantationen das fremde Genom sogar irreversibel denaturiert werden kann (Literatur bei Jinks 1967). Da weiter das Cytoplasma die Kerngröße und den zeitlichen Ablauf der DNA-Synthese beeinflusst (vgl. Maclean 1977) ja die DNA-Synthese selbst von der plasmatischen Konstitution abhängt (s.o.), muss es auch einen Zusammenhang zwischen dem Plasmon und spezifischer Replikation von Genen geben.
[Vielleicht hängt diese Frage auch mit dem zweiten genetischen Code zusammen. Vgl. C. de Duve (1988): The second genetic code. Nature 333, 117-118. Literatur zum Thema Promotersequenzen und Transkriptionsfaktoren siehe zum Beispiel S. L. Wolfe (1993): Molecular and Cellular Biology. Belmont, Cal. S. F. Gilbert (1991): Developmental Biology. 3. Aufl. Sunderland, Mass.]
Gurdon stellt 1974, p. 111 fest (ähnlich De Robertis und Gurdon 1980/1985):
We conclude from the nuclear transfer experiments summarized that egg cytoplasm contains an inducer of DNA synthesis and a repressor of RNA synthesis which operate independently on different classes of genes. Attempts to identify these components are very greatly facilitated by the fact that they are absent or inactive in oocytes which are similar to eggs in so many respects.
Bei Ascaris finden wir darüber hinaus auch eine plasmatisch kontrollierte Chromosomenverkleinerung und Chromosomenabbau, ebenso bei cecidomyiden Fliegen (Information zum Teil zumindest wieder aus dem Kern?).
- Wie das alles im Einzelnen vorsichgeht, ist molekularbiologisch noch weitgehend ungeklärt, so dass eine endgültige Aussage zu Lamprechts Progen-Konzept noch verfrüht erscheint. (Die Reguation von Splicing und Processing-Vorgängen sei hier noch erwähnt: Für das Splicing der Vorläufer-m-RNA als Transkriptionsprodukt von Mosaikgenen bedarf es spezifischer Enzyme, deren Struktur wiederum von anderen Genen festgelegt wird. Diese 'Progene' sind jedoch nicht für jedes einzelne Gen spezifisch verschieden, so dass etwa jedem Strukturgen ein Splicing-Gen zugeordnet wäre.)
Zur Funktion der Gene haben wir inzwischen gelernt, dass Regulatorgene für das Ein- und Abschalten ganzer Genbatterien zuständig sind. Der Gedanke, dass jedem Strukturgen (genau) ein Progen für dessen Funktion und Replikation zugeordnet sei, ist aus dieser Sicht wenig wahrscheinlich. (Siehe jedoch S. 13.)
Ich habe diese verschiedenen Punkte hier aufgeführt, weil sie die Hierarchie von Genregulations- und Replikationsmechanismen in Zusammenarbeit mit dem Plasma veranschaulichen, die den Lamprechtschen Ansatz zumindest insofern bestätigen, als nicht jedes Gen für sich allein schaltet und waltet und sich selbst repliziert (wie das zu Lamprechts Zeit häufig postuliert wurde und darüber hinaus noch bis in die Gegenwart von Dawkins und anderen vertreten wird).
["Protein synthesis is an integral part of the overall pathway of gene expression. Gene expression can be controlled at essentially all levels of the process: through gene rearrangements; at the levels of transcription, mRNA processing, transport to the cytoplasm, and stability; at the level of translation; and through posttranslational modifications."
"...With weIl over 100 macromolecular components, protein synthesis can probably claim to be the most complex multienzyme system studied at the molecular level." (J. W. B. Hershey, R. Duncan, and M. B. Mathews 1986, pp. 1 und 13; in: Translational control. Ed.: M. B. Mathews. Cold Spring Harbor Laboratory.
J. Brachet 1987, p. 309 nach Diskussion neuerer Forschungsergebnisse zur Genregulation und Beispielen zur cytoplasmatisch-translationalen Kontrolle pp. 295, 296 und 298: "...many basic problems remain unsolved. Several young and bright molecular biologists have asked me the same question, "What shall we do when all the genes have been cloned and sequenced?" I do not think that a complete knowledge of the mouse genome will be enough to understand fully how a mouse egg gives rise to a mouse." (Nucleocytoplasmatic interactions in morphogenesis. International Review of Cytology 100, 249 - 318.)]
Als weitere Schwierigkeit für das Lamprechtsche Konzept sei daran erinnert, dass nach den Untersuchungen Ohnos schon in den Hybriden zahlreiche Gene des väterlichen Genoms gar nicht erst "angeschaltet" werden - das Vorhandensein und das Fehlen der Gene und "Progene" wurde von Lamprecht jedoch aus der Genfunktion erschlossen (vgl. auch Lamprecht 1948, pp. 134/135 und 1974, p.564). Das letztere kann zwar zutreffen (vgl. z.B. Tunner und Heppich 1981 zum Thema PREMEIOTIC GENOME EXCLUSION DURING OOGENESIS IN THE COMMON EDIBLE FROG, RANA ESCULENTA; vgl. auch Heppich et al. 1982), muss aber nicht, wie die Replikation von Pseudogenen, Transposons und repetitiven Genen in der Meiose zeigt.
Es bleibt auch unverständlich, dass die "Progene" immer pünktlich bei den ersten Meiosen verbraucht sein sollen (vgl. Lamprecht 1948, p. 136 zum Verbrauch der "Progenmasse"), während doch die Teilung der Somazellen auch bei den von Lamprecht untersuchten Blütenpflanzen noch eine geraume Zeit weitergehen kann (bei Bäumen sogar über Jahrzehnte hinaus, - bei Tieren laufen ebenfalls beide Prozesse gleichzeitig ab). Auf der anderen Seite ist klar, dass die in der Eizelle produzierten morphogenetischen Determinanten in der Morphogenese verbraucht werden - langlebige mRNA z.B. - und für die nächste Generation neu synthetisiert werden müssen.
Ein weiterer Einwand betrifft die Gleichsetzung ein Merkmal = ein Gen in den obigen Lamprechtschen Ausführungen. Mendelsche Spaltung kann zwar auf einen einzigen Faktor zurückzuführen sein (z.B. beim Ausfall der Farbstoffsynthese durch ein Grundgen für die Blütenfarbe), aber für die volle Farbstoffsynthese sind zahlreiche Gene notwendig. Dooner und Nelson bemerken 1979, p. 313:
The genetic control of anthocyanin pigmentation in the aleurone layer of the maize kernel has long been regarded as a textbook example of genic interaction. The interplay of many genes leads to the formation of the final product. Mutation in at least eight of these genes (A, A2, Bz, Bz2, C, C2, R and Vp) results in failure of the aleurone to produce anthocyanin.
(Vgl. auch Dooner 1983, p. 140, wo er - inzwischen - von mindestens 9 notwendigen Loci spricht.)
Lamprecht hat ein solches Gensystem übrigens detailliert für die Blütenfarben von Pisum beschrieben (vgl. pp. 194-194), so dass seine grundsätzliche Gleichsetzung ein Gen = ein Merkmal im Zusammenhang mit dem interspezifischen Gensystem nicht ganz verständlich ist.
Die Kollagensynthese z. B. verlangt "a dozen or more enzymes" (Pain 1987, p. 298), und die Nitrogenase ist "an extremely complex enzyme, produced by 17 - 20 nif genes" (McCormick 1988, p. 383) (beides Reviews).
Vollmert beschreibt (1983, p. 32) die allgemeine Situation treffend wie folgt:
Bis zu 10 oder 12 neue Gene (entsprechend 10 oder 12 neuen Enzymen) müssen entstehen, damit auch nur die Synthese eines einzigen neuen Stoffes (z.B. im Rahmen der Photosynthese oder der Atmungskette oder der Blutregenerierung) ermöglicht wird.
(p. 33:) Da ein neuer Stoff in aller Regel nur im Rahmen eines größeren Verbundes von 20 bis 30 anderen Stoffen von Nutzen ist, kann eine neue Eigenschaft [Im Sinne einer Neuorganisation für höhere systematische Unterschiede (Anmerkung von mir).] erst nach der Entstehung von 20 mal 10 = 200 neuen Genen oder (im Durchschnitt) 200 mal 1500 = 300 000 neuen Nukleotidsequenzen in Erscheinung treten...
Artentrennende Merkmale, d.h. Merkmale, die nicht von der einen Art in die andere und vice versa überführt und in fertilen Nachkommen erhalten werden können, dürften demnach häufig Genwirkketten mit Regulatorsystemen voraussetzen.
Der Ausfall von Regulatorgenen kann zwar ganze Genwirkketten abschalten - aber Mutationen in diesen Genen können keine Genwirkketten aufbauen (das gleiche trifft auf Promotorsequenzen zu). Insofern scheinen Lamprechts Exmutanten auch nicht mit primären Artdifferenzen vergleichbar zu sein. Aber das letzte Wort ist hierüber noch nicht gesprochen, da derzeit zahlreiche Autoren Regulationsprozesse als Hauptfaktor der Entstehung neuer Arten postulieren.
Adams und Rinne begründen ihre Auffassung wie folgt (1982, p. 312):
Patterns of gene regulation are possibly central to evolutionary change since there is a remarkable similarity of structural gene function between simple and complex organisms (Britten and Davidson, 1971). About 93 % of the enzyme activities listed in the International Enzyme Commission tables are known to be present in both prokaryotes and mammals. Therefore evolutionary events leading to the development of different organisms are unlikely to be the product of numerous mutations in the spectrum of enzymes produced by different organisms.
(Vgl. auch King und Wilson 1975, Stanley 1979, Hedrick und McDonald 1980, Sybenga 1981, Adams et al. 1982, Raff und Kaufman 1983 u.v.a.)
Andererseits sollte man bei dieser Frage berücksichtigen, dass auch die zu postulierenden zahlreichen Regulationsprozesse selbst - beispielsweise für den Aufbau neuer Organe (abgesehen von den für die Organfunktion sowieso notwendigen neuen Stoffen) - das harmonische Zusammenspiel zahlreicher Gene notwendig macht, so dass auch in diesem Falle viele Strukturen der Organismenwelt (die Arten kennzeichnen und durch ihre Nicht-Überführbarkeit voneinander trennen) nicht allein mit einzelnen interspezifischen Genen zu erklären sind.
Anders kann es sich jedoch mit Lamprechts Beispielen der Kotyledonenstellung und dem Herablaufen der Narbe auf der Innen- bzw. Außenseite der Griffelspirale bei Phaseolus verhalten. Diese (im Grunde genommen) geringfügigen Unterschiede ließen sich im wesentlichen vielleicht mit zwei Regulatorgenpaaren erklären, die nicht reziprok von der einen Linie in die andere überführbar sind.
Wall und York bemerken jedoch schon 1957, p. 71:
Seedlings of Phaseolus have two distinct cotyledon positions. In P. coccineus the cotyledons are hypogaeous while in P. vulgaris they are epigaeous. The position of cotyledons in the seedling stage has long been considered a valid taxonomic character in this genus. However, Freytag reports races of P. coccineus, found in Mexico in the states of Puebla, Oaxaca, and Chiapas, which bear their cotyledons half-way between the first leaves and ground level, i.e. epigaeously. He also states that in this same region are found races of P. vulgaris which have their cotyledons practically touching the ground. This report leads to the inference that interspecific hybridization has occurred between these species.
Die Autoren sind weiter der Auffassung, dass hier "quantitative factor differerces" eine Rolle spielen. Sie fanden allerdings in ihrer F2, F3 und F4 das gleiche Ergebnis wie Lamprecht und schreiben (p. 73):
It is suggested that the explanation for the skewing of the F2 in the direction of the vulgaris parent is the result of the elimination of gametes carrying high proportions of genes of P. coccineus and/or zygotic elimination of zygotes resulting from the union of gametes with a high proportion of coccineus genes. In this manner coccineus genes would be eliminated from the gene complex. It is to be remembered that we are dealing with coccineus genes functioning in vulgaris plasma and that they may therefore be at a selective disadvantage. The progressive shift of the F3 and F4 toward the vulgaris parental type is consistent with such an interpretation.
(Vgl. auch Haq et al. 1980 und Smartt 1981.)
Wir dürfen bei den verschiedenen Einwänden und Überlegungen nicht aus den Augen verlieren, dass nach allen bisherigen Untersuchungen die empirischen Befunde Lamprechts korrekt sind und einer Erklärung bedürfen: bestimmte Merkmale sind bei Kreuzung verschiedener Phaseolus- und Chrysanthemum-Linien nicht reziprok überführbar. Damit stimmen die Befunde Ohnos an Forellen- und Wachtelhybriden im Prinzip überein sowie die oben ausführlich dokumentierte Genregulation durch das Ei-Plasma bzw. die Bedeutung des Plasmas überhaupt. (Siehe zur präferentiellen Aktivierung von maternellen Allelen in Arthybriden weiter Frankel 1983 und Parker et al. 1985.)
Wir müssen diese Fakten ganz klar von den Schwächen der prämolekularen Genvorstellungen Lamprechts, insbesondere seines Progen-Konzepts, trennen, da wir sonst für den Artbegriff entscheidende Funde mit Lamprechtschen Fehlern über Bord werfen würden.
Ich möchte weiter festhalten, dass bei den zur Diskussion stehenden Fakten die Genregulation über die Konstitution des Ei-Plasmas mit seinen morphogenetischen Determinanten (s.o.) und die Meiosis die entscheidenden Rollen spielen. Das hatte auch Lamprecht klar erkannt.
Bestimmte Merkmale verschiedener Organismen sind aufgrund dieser (mit Sicherheit ineinandergreifenden) Regulationssysteme, die zugleich Barrieren gegen die Expression und Repikation andersartiger Gene und Genwirkketten (samt Regulatorgenen und Promotoren) bilden, nicht reziprok überführbar.
Wir wissen jedoch derzeit noch zu wenig, um diesen festen Rahmen mit molekulargenetischen Details zu füllen ("...most of the crucial causal links between genome and embryo remain largely unknown." Davidson 1986, p. 2)
Horvitz schreibt 1980, p. 435:
The relationship between genes and development is unknown. It appears that development and differentiation are based on differential gene activity, and considerable effort has been dedicated to determining the mechanism used in elicitating this differential gene activity (e.g. Gurdon 1974; Davidson 1976). Nonetheless, little is understood about the genetic control of gene expression during development.
Trotz einiger überraschender neuer Befunde (vgl. Gehring 1985; einleitende Darstellung bei Hogan 1985), die zu Hoffnung Anlass geben, hat sich in den letzten Jahren die Situation noch nicht grundlegend geändert.
Ghysen, Jan und Jan weisen in der Einleitung zu ihrer Arbeit über die Determination der Segmente im Zentralnervensystem von Drosophila ebenfalls auf diesen Punkt hin, wenn sie (1985, p. 943) schreiben:
How genes control the development of an organism is a major unanswered question in biology. Some cues about the control are emerging in one case, the process of segmentation in Drosophila. Segmentation is an essential step in the development of all higher organisms.
["The recent isolation by Dolecki et al. of a homeobox gene from sea urchin, which lacks a segmented body plan, forces a reconsideration of the possibility that homeoboxes tag genes involved in the specification of segmental information. ...The big question 'what do they do' still remains." (R. L. Chisholm 1986: Homeoboxes: what do they do? TIG 2, 224 - 225) - (Vgl. auch Cell 55, 537-540; 1988) Siehe jedoch De Robertis et al.: Spektrum der Wiss. Sept. 1990, p. 84 ff.; Sommer et al. EMBO J. 9, 605-613.]
( Präziser müsste es heißen: "How genes are controlled in the development of an organism...") Sieht man sich solche Arbeiten genauer an, so stellt man meist fest, dass sie wenig über diesen Punkt zu sagen haben. Statt dessen wird die Bedeutung einzelner Gene bzw. spezifischer DNA-Sequenzen für bestimmte Entwicklungsprozesse herausgearbeitet (was selbstverständlich eine wichtige Aufgabe ist). Über die Regulation heißt es in der Arbeit von Ghysen et al. nur:
(p. 943:) The segmental determination of all segments posterior to the mesothorax depends on a single gene cluster, the bithorax complex (BX-C; ...).
(p. 946:) The expression of the BX-C is controlled by various regulatory genes (..lit.). One of them, Rg-bx, is required for the normal expression of the complex (..lit.). A leaky allele of Rg-bx, named trx, is homozygous viable (Ingham, 1981).
...The expression of trx is increased in zygotes derived from trx/Df-red mothers where Df-red deletes the trx gene (Ingham and Whittle, 1980). This maternal effect is also observed at the level of the CNS [central nervous system] (Figures) without any parallel effect on the epidermis.
Genaueres ist derzeit (1986) zur Frage, wie nun die Regulatorgene im vorliegende Falle ihrerseits reguliert werden, kaum bekannt [Daten bis Anfang 1988 vgl. Nachtrag p. 534.]. Interessant ist jedoch, dass auch hier eine maternelle Wirkung beobachtet werden konnte. Ingham und Whittle weisen darauf hin (1980, p. 613), dass das trx Gen sowohl während der Oogenese als auch der Embryogenese transkribiert wird, wodurch oder "von wem" es selbst reguliert wird, aber erfährt man nicht (vgl. auch Ingham 1985, Jeffery 1985). Hier liegen noch viele offene Probleme.
Wir wollen jedoch festhalten, dass wir hiermit einen Fall vorzuliegen haben, in dem ein für die Ontogenese von Drosophila wichtiges Regulatorgen bereits in der Oogenese transkribiert wird (weitere Beispiele bei Anderson et al. 1985 a und 1985 b, Rebagliati et al. 1985).
[Davidson 1986, p. 109: "A conclusion of general significance drawn from studies on maternal mutants as a class is that the expression of a majority of all Drosophila genes that can mutate to lethality is required during oogenesis." (Vom Verfasser kursiv.) - Was die Möglichkeit sekundärer Artbildung (vgl. vorliegende Seite Mitte) wieder begrenzt.]
Es ist klar, dass 'artfremde Gene' (bei Überführung in das Plasma einer anderen Art) vom mütterlichen PLasma nicht "angeschaltet" werden können, wenn es keine für sie passenden Regulatortranskripte (m-RNA) und -Proteine gibt, die ja in vielen Fällen bereits für den normalen Start der Entwicklung in ausreichender Menge vorhanden sein müssen. Die mit dem Chromosomensatz des Vaters gelieferten Regulatorgene kommen 1. häufig zu spät und 2. müssen diese Regulatorgene selbst ja auch wieder reguliert werden. Ist das mütterliche Plasma nicht darauf eingestellt, dann wird der ganze Komplex gar nicht erst in Betrieb genommen.
Den nächsten Schritt würden Regulatorgene bilden, die nur (oder zumindest vorwiegend) in der Oogenese transkribiert werden. In diesem Falle kommen die vom Vater gelieferten Regulatorgene grundsätzlich zu spät.
Als weiterer wichtiger Punkt sei hervorgehoben, dass eine solche Artbarriere auch sekundären Ursprungs sein könnte. Nehmen wir bei zwei geographisch oder sonstwie isolierten Populationen den Ausfall unterschiedlicher Regulator- (und zugeordneter Ziel-) Sequenzen an. In der Oogenese werden demnach auch nicht mehr die entsprechenden Transkripte und stabilen m-RNAs synthetisiert. Resultat: bestimmte Merkmale nahverwandter Arten folgen bei Kreuzung nicht mehr den Mendelschen Regeln und können damit auch nicht mehr von der einen Linie in die andere (und reziprok) überführt werden (vgl. dazu die Forellen- und Wachtelbeispiele von Ohno zitiert pp. 223-224).
Offen bleibt dabei immer noch die Frage nach der Regulation der Regulatorgene.
Lamprechts Bohnenbeispiel gibt uns zu dieser Frage einen Hinweis.
Versuchen wir zunächst einmal die nicht überführbaren Merkmale allein mit Regulatorgenen im obigen Sinne zu erklären: Die ursprüngliche Fähigkeit zur Stengelbildung unterhalb der Kotyledonen sei bei Phaseolus coccineus durch Mutation eines Regulatorgens verlorengegangen, bei Ph. vulgaris hingegen werde diese Stengelbildung durch ein sowohl in der Oogenese als auch in der Ontogenese transkribiertes, voll funktionsfähiges Regulatorgen ermöglicht.
Bei Kreuzung der beiden Linien ist die F1 intermediär, weil in der Ontogenese durch Ausfall (oder starke Veränderung) des Regulatorgens von coccineus der in diesem Gen hemizygote Zustand vorliegt. Man würde jedoch einen reziproken Unterschied erwarten, da vulgaris mit der bereits in der Oogenese begonnenen Transkription über eine größere Anzahl von Transkripten und stabilen m-RNA verfügen sollte als bei coccineus als Mutter. In der F1 kann dann durch das funktionsfähige Allel von vulgaris wieder die für die Expression der epigäischen Keimblattstellung notwendige m-RNA in der Oogenese synthetisiert werden. Durch eine solche Prädetermination würde das coccineus-Allel (auch bei Homozygotie) in der F2 nicht exprimiert werden. Allerdings müsste dann in zunehmendem Maße von der F3 an das coccineus-Allel auftreten, da das Viertel der in diesem Allel homozygoten F2 Rekombinanten diese m-RNA nicht mehr produzieren kann. Das Gegenteil ist der Fall! Die coccineus-Merkmale treten überhaupt nicht auf und die vulgaris-Merkmale setzen sich im vermehrten Maße durch (vgl. die oben zitierten Arbeiten von Lamprecht und Wall und York).
Machen wir den gleichen Ansatz mit coccineus als Mutter, dann sieht die Situation folgendermaßen aus: Wir haben zwar keine Produktion von funktionsfähiger m-RNA in der Oogenese, aber die F1 ist intermediär, da durch das intakte vulgaris-Allel genügend Transkripte für die benötigten m-RNA-Moleküle bereitgestellt werden. Bei den gleichen Annahmen wie bei vulgaris als Mutter dürften nun in der F2 ebenfalls keine coccineus-Pflanzen auftreten, doch das Viertel der bereits in der F2 im coccineus-Allel homozygoten Pflanzen müsste dann in der F3 nur coccineus-Phänotypen hervorbringen (vgl. dazu das Kapitel von Kühn und Hess über Prädetermination 1984, p. 255 ff.). Das alles trifft nicht zu! Wir müssen feststellen, dass wir mit der Funktion von Regulatorgenen allein als Erklärung für die Genetik der Keimblattstellung der beiden Bohnenlinien nicht auskommen (auch nicht bei Einbeziehung des Prädeterminationsphänomens).
Ähnlich Lamprecht komme ich zu dem Schluss, dass in der Meiose der F1-Pflanzen etwas geschieht, was die Replikation und/oder Expression und Rekombination der oben behandelten Allele im 'artfremden' Plasma nicht mehr zulässt; - d.h. die (angenommenen) Regulatorgene selbst müssen von Plasmafaktoren kontrolliert werden.
Es muss sich dabei um ein vom Kern unabhängiges Kontrollsystem im Plasma handeln, denn diese Kontrolle geht ausschließlich von der Mutterpflanze aus. (Lamprecht hat das übrigens mit der Fremdplasma-Anreicherung und dem darauf folgenden Umschlag von der einen 'Art' in die andere durch Plasmaentmischung überzeugend veranschaulicht - vgl. Lamprecht 1966.)
Tatsächlich ist das Bohnenbeispiel hervorragend geeignet, um die meiotisch-plasmatische Kontrolle bestimmter Gene zu belegen. Diese plasmatische Kontrolle besteht im vorliegenden Fall in der totalen Blockade der 'art-fremden' Allele.
Mit diesen Befunden stimmen jedenfalls die irreversiblen Veränderungen überein, die die Kerne bei Transplantation in artfremdes Plasma erleiden (Zusammenfassung z.B. bei Jinks 1967, Beale und Knowles 1978, Gurdon 1986).
Wie weit lassen sich diese Befunde verallgemeinern? Gibt es ein plasmatisches Kontrollsystem für einen entscheidenden Teil (zumindest) der Regulatorfunktionen im Pflanzen- und Tierreich?
Für den Artbegriff ist das eine entscheidende Frage; denn zahlreiche Befunde aus dem Bereich der Chloroplasten und Mitochondrien-Genetik (den ich jedoch nicht mit dem obigen plasmatischen Kontrollsystem identifizieren möchte, da ja auch Chloroplasten und Mitochondrien-Genom reguliert und 'kontrolliert' werden müssen) deuten bereits an, dass ein solches (meiotisch-) plasmatisches Kontrollsystem wesentlich konstanter (besser "gepuffert") sein dürfte als das Kemsystem mit seinen zahlreichen Mutationen und Instabilitäten.
Zur Frage, was die Ursachen der Plasmonstabilität sein könnten (dabei geht es vor allem um das Chloroplasten- und Mitochondrien-Genom), schreibt Grun 1976, p. 336 unter anderem:
In organisms having plasmon factors transmitted only through the female, no new genetic plasmon factors would be introduced into the egg and therefore no genetic recombination of plasmon genes could occur. Once the process of mitotic sorting-out had established a homoplasmon (Chapter 2), that type would be maintained by a regular copying process; the only variability introduced would come about through the infrequent occurrence of mutations.
Dabei ist noch zu berücksichtigen, dass das Chloroplasten- und Mitochondrien-Genom auch in den einzelnen Organellen in jeweils zahlreichen Kopien vorliegt. Mutationen in diesem Bereich müssen sich sowohl gegen die Übermacht der zahlreichen Kopien der Ausgangssequenzen durchsetzen als auch in natürlichen Populationen einen klaren Selektionsvorteil haben. In zahllosen Mutationsexperimenten mit Pflanzen tritt regelmäßig ein stattlicher Anteil kernbedingter Chlorophyllmutanten auf, Plastidenmutanten sind dagegen ausgesprochen selten.
Bei der Frage nach einem Selektionsvorteil muss man sich noch folgende Aussage Kimuras (1983, p. 35) für die Kernmutationen vor Augen halten:
Although a large number of mutants arise in each generation in any reasonably large population, the majority are lost by chance within a small number of generations (Fisher, 1930b; Kimura and Ohta 1969b). It is often not realized that this is true not only for deleterious and selectively neutral mutants but also for advantageous mutants unless the advantage is very large. For example, if a mutant has a 1 % selective advantage, the chance is only about 2 % that it will eventually spread through the whole population (Haldane, 1927; Fisher 1930b). In the remaining 98 % of the cases it will be lost by chance from the population without being used in evolution.
Soweit einige Überlegungen zur Frage nach der Konstanz der bekannten Plasmonfaktoren. Die im Vergleich zu den Kerngenen wesentlich größere Stabilität des Plasmons war auch eine der Ursachen für ihre verspätete systematische Erforschung und verzögerte Anerkennung als autonomes genetisches System (vgl. Michaelis zitiert p. 214) - obwohl Correns bereits anfang unseres Jahrhunderts klare Befunde zu diesem Thema vorgelegt hatte.
Könnte es sich bei dem von Lamprecht, Dillon, Blechschmidt u.a. postulierten plasmatischen Kontrollsystem nicht um einen ähnlichen Vorgang handeln? Denn die bisher vorliegenden Befunde deuten auf ein im Vergleich zu allen bisher bekannten genetischen Systemen noch stabileres Regulationssystem, speziell durch die meiotisch-plasmatische Kontrollfunktion 'interspezifischer' Gene.
________________________
NACHTRAG ZU SEITE 236 (REGULATION DER REGULATORGENE)
M. P. Scott und S. B. Carrol fassen in ihrem Review-Artikel THE SEGMENTATION AND HOMEOTIC GENE NETWORK IN EARLY DROSOPHILA DEVELOPMENT (Cell 51, 689 - 698; 4. Dezember 1987) die hauptsächlichen Ergebnisse zu diesem Thema geschickt zusammen. Ein paar für unsere Fragestellung wichtigen Punkte seien im folgenden zitiert (p. 689):
Embryonic pattern formation begins during oogenesis, when the oocyte is packed with maternally encoded mRNAs and proteins. Some of the maternally active genes are segmentation genes, as revealed by maternal-effect mutations that alter the segmentation pattern in the progeny of homozygous mutant female flies (lit...).
P. 690: Although the formal regulatory relationships among the genes are becoming clear, the mechanisms involved are still mysterious. It is not known how the gene interactions result in patterns such as stripes and, in particular, whether the interactions are direct or mediated through other genes. The mechanisms must be both precise, in that adjacent cells must in some cases behave very differently, and flexible, in that the number and exact arrangement of cells is different in different embryos. The mechanisms cannot work through counting cells, since embryos with drastically different numbers of nuclei can still form stripes of the normal size (Sullivan 1987). Furthermore, he simplicity of he striped patterns conceals regulatory subtlety. Different stripes depend on different sets of upstream functions; not au ftz stripes, for example, require the same gap gene function.
P.696: .....we expect that the most provocative information will come from the analysis of the expression and function of maternal gene products, the identification of the molecular mechanisms controlling interactions between the different tiers of the zygotic gene hierarchy, and the characterization of cis-acting elements of pattern-regulating genes that respond to specific spatial, temporal, and tissue-specific regulatory proteins.
Dem an weiteren Details interessierten Leser sei dieser Artikel sowie die folgenden empfohlen:
Weeks, D.L. and D.A. Melton (1987): A MATERNAL mRNA LOCALIZED TO THE VEGETAL HEMISPHERE IN XENOPUS EGGS CODES FOR A GROWTH FACTOR RELATED TO TGF-ß. Cell 51, 861 - 867
Ingham, P.W., N.E. Baker and A. Martinez-Arias (1988): REGULATION OF SEGMENT POLARITY GENES IN THE DROSOPHILA BLASTODERM BY FUSHI TARAZU AND EVEN SKIPPED. Nature 331, 73 - 75